

Copyrights: Gopikrishna Agraharam, Agnishwar Girigoswami, Pemula Gowtham, Koyeli Girigoswami, 2023. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
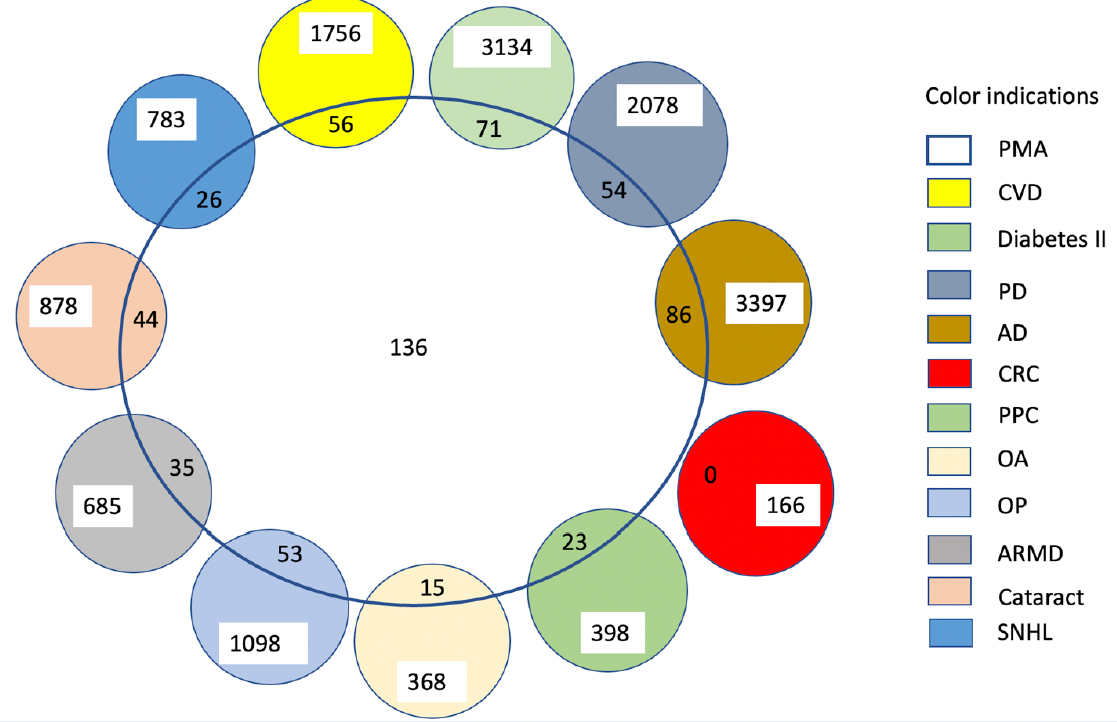
Aging is an irreversible biological process observed in living organisms, with each cell demonstrating this mechanism. Age-related decline in cellular integrity due to various endogenous and exogenous factors and mechanisms contributes to several diseases based on the site or location of the decline. Cardiovascular diseases, diabetes, cancer, osteoarthritis, and osteoporosis are some examples of age-related diseases. Reports suggest several premature aging genes contribute to the genetic, pathological, and physiological variations of an organism that is similar to the aging organism and might explain the genes associated with aging. We used the DisGeNET web tool to retrieve genes associated with premature aging. We used the Cytoscape software and STRING online tool for identifying protein-protein networks. We retrieved 136 genes associated with premature aging and compared their association with age-related diseases. Many of these premature agingassociated genes were associated with cardiovascular disease (56), diabetes type II (71), Parkinson's disease (54), Alzheimer's disease (86), prostate cancer progression (23), osteoarthritis (15), osteoporosis (53), age-related macular degeneration (35), cataracts (44), and sensorineural hearing loss (26). This article provides a brief review of some of the genes involved in premature aging.
Introduction
Aging is a chronological process that is caused by damage to biomolecules, such as DNA, RNA, proteins, and cellular organelles. This damage leads to changes in the functions of cellular organelles, such as the mitochondria, lysosomes, and endoplasmic reticulum. This results in a variety of multicellular functions and age-related diseases that result in death1, 2, 3, 4. This damage is primarily caused by oxidative DNA stress, which results in single-stranded breaks, double-stranded breaks, and modifications to nitrogen bases or sugar-phosphate backbones5, 6. Mitochondria are the powerhouse of the cell and generate energy via the electron transport chain (ETC), which releases free radicals, such as O2- and H+, into the inner mitochondrial membrane space and outer mitochondrial space. These radicals damage cellular DNA, RNA, proteins, and mtDNA, thereby damaging the mitochondria, which results in decreased cellular energy production7. Lysosomes protect cells by degrading damaged or misfolded proteins and malfunctioning cytoplasmic macromolecules via autophagy. Mutations of lysosomal proteins and enzymes lead to lysosomal malfunctioning, which causes excessive amounts of damaged biomolecules to accumulate inside a cell, causing cell damage8. Modifications in rRNA and r-proteins due to oxidative stress leads to ribosomal dysfunction, meaning newly translated proteins may not be able to effectively perform their activities in the cell9.
The past five decades of research indicate that chronic damage or genetic mutations in some of the genes involved in the aging process leads to physiological and pathological changes that are similar to those seen in the aging process10. Bioinformatic tools provide advantages in retrieving enormous amounts of data, while network biology tools can identify associations between genes and proteins. Gene silencing plays a role in inhibiting colon cancer11. In this mini-review, we identified 136 genes associated with premature aging using DisGeNET12, a disease-gene association finding tool, and identified which genes were also associated with other age-related diseases, such as cardiovascular disease, osteoporosis, osteoarthritis, and cataracts. Here, we have briefly reviewed several genes associated with premature aging.
Methodology
We used the DisGeNET12 database for finding genes associated with premature aging and age-related diseases and the STRING database13 and Cytoscape software14 for identifying protein-protein interactions for the genes associated with premature aging. The similarity search was done manually, and relevant articles related to the genes were found using the Google Scholar, Scopus, and PubMed databases.
Aging genes and similarities
We identified 136 premature aging genes (premature aging syndrome; CUI: C0231341) using DisGeNET. Those with the highest scores were Klotho (KL), Werner syndrome gene (WRN), excision repair cross complementation group 6 (ERCC6), amyloid-beta precursor protein (APP), exostosin glycosyltransferase 1 (EXT1), lamin A (LMNA), RecQ like helicase 3 (RECQL3/BLM), RecQ like helicase 4 (RECQL4), tumor protein p53 (TP53), and Sirtuin 1 (SIRT1). When we manually compared the premature aging-associated genes with those associated with age-related diseases, we found several commonalities: 56 of the 1756 cardiovascular disease-associated genes (Cardiovascular Diseases; CUI: C0007222), 71 of the 3134 diabetes type II-associated genes (Diabetes Mellitus, Non-Insulin-Dependent; CUI: C0011860), 54 of the 2078 Parkinson’s disease-associated genes (Parkinson Disease; CUI: C0030567), 86 of the 3397 Alzheimer’s disease-associated genes (Alzheimer’s Disease; CUI: C0002395), 0 of the 166 colorectal cancer-associated genes (Susceptibility to Colorectal Cancer, 12; CUI: C3554460), 23 of the 398 prostate cancer-associated genes (Prostate Cancer Progression; CUI: C1739135), 15 of the 368 osteoarthritis-associated genes (Osteoarthritis, Knee; CUI: C0409959), 53 of the 1098 osteoporosis-associated genes (Osteoporosis; CUI: C0029456), 44 of the 878 cataract-associated genes (Cataract; CUI: C0086543), 35 of the 685 macular degeneration-associated genes (Age-related Macular Degeneration; CUI: C0242383), 26 of the 783 hearing loss-associated genes (Sensorineural Hearing Loss [disorder]; CUI: C0018784)12. As colorectal cancer was not associated with any premature aging genes, it has been excluded from Table 1 (Table 1, Figure 1).

| S/No | PMA | CVD | Diabetes -type II | PD | AD | PPC | OA | OP | ARMD | Cataract | SNHL | No. of age-related diseases the genes are present |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | KL | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 2. | WRN | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 3. | ERCC6 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 4. | APP | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 5. | EXT1 | ✓ | ✓ | 2 | ||||||||
| 6. | LMNA | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 7. | BLM | ✓ | ✓ | ✓ | 3 | |||||||
| 8. | RECQL4 | ✓ | ✓ | 2 | ||||||||
| 9. | TP53 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 9 | |
| 10. | SIRT1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 11. | ZMPSTE24 | ✓ | ✓ | ✓ | 3 | |||||||
| 12. | CISD2 | ✓ | ✓ | ✓ | 3 | |||||||
| 13. | ATM | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 14. | FGF23 | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 15. | HTRA2 | ✓ | ✓ | ✓ | 3 | |||||||
| 16. | HSPA9 | ✓ | ✓ | ✓ | 3 | |||||||
| 17. | SPRTN | 0 | ||||||||||
| 18. | ARNTL | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 19. | CDKN2A | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 20. | IGF1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 10 |
| 21. | CSH1 | 0 | ||||||||||
| 22. | CSH2 | 0 | ||||||||||
| 23. | SOX2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 24. | ROBO3 | ✓ | 1 | |||||||||
| 25. | SOD1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 26. | BUB1B | ✓ | 1 | |||||||||
| 27. | ANGPTL2 | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 28. | VDR | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 29. | BMI1 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 30. | TH tyrosine hydroxylase | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 31. | ELN | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 32. | ERCC8 | ✓ | ✓ | 2 | ||||||||
| 33. | CYP27A1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 34. | DKC1 | ✓ | ✓ | 2 | ||||||||
| 35. | EMD | 0 | ||||||||||
| 36. | CDKN1A | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 37. | ERCC2 | ✓ | ✓ | ✓ | 3 | |||||||
| 38. | PYCR1 | ✓ | 1 | |||||||||
| 39. | SFRP1 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 40. | CHMP1B | 0 | ||||||||||
| 41. | WRNIP1 | 0 | ||||||||||
| 42. | SOD2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 43. | SOD3 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 44. | TWNK | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 45. | SNAI2 | ✓ | 1 | |||||||||
| 46. | PSMD2 | ✓ | ✓ | 2 | ||||||||
| 47. | TNMD | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 48. | RAD51 | ✓ | ✓ | ✓ | 3 | |||||||
| 49. | RBBP4 | 0 | ||||||||||
| 50. | BCL2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 51. | SMURF2 | ✓ | 1 | |||||||||
| 52. | SCT | ✓ | 1 | |||||||||
| 53. | SLC3A2 | 0 | ||||||||||
| 54. | SRSF5 | ✓ | ✓ | 2 | ||||||||
| 55. | BRCA1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 56. | CAT | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 57. | IKBKG | ✓ | 1 | |||||||||
| 58. | CAV1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 59. | RUNX2 | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 60. | TP63 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 61. | BANF1 | ✓ | ✓ | 2 | ||||||||
| 62. | RECQL5 | 0 | ||||||||||
| 63. | ATG5 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 64. | TBPL1 | ✓ | ✓ | ✓ | 3 | |||||||
| 65. | CLOCK | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 66. | CUL4A | ✓ | 1 | |||||||||
| 67. | SPNS1 | 0 | ||||||||||
| 68. | CASP2 | ✓ | 1 | |||||||||
| 69. | SRF | ✓ | ✓ | 2 | ||||||||
| 70. | ROPN1L | ✓ | 1 | |||||||||
| 71. | TERC | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 72. | TERF2 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 73. | TFAM | ✓ | ✓ | ✓ | 3 | |||||||
| 74. | TOP3A | ✓ | 1 | |||||||||
| 75. | C3 complement C3 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 76. | VCP | ✓ | ✓ | ✓ | 3 | |||||||
| 77. | XPO1 | 0 | ||||||||||
| 78. | EDS8 | 0 | ||||||||||
| 79. | NCOR2 | ✓ | 1 | |||||||||
| 80. | MAPK1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 81. | CFDP1 | ✓ | ✓ | 2 | ||||||||
| 82. | EFEMP1 | ✓ | ✓ | 2 | ||||||||
| 83. | FGF1 | ✓ | ✓ | ✓ | 3 | |||||||
| 84. | SIRT2 | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 85. | FOXO3 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 86. | EXOSC2 | ✓ | 1 | |||||||||
| 87. | COMMD3-BMI1 | ✓ | ✓ | 2 | ||||||||
| 88. | KCNH4 | ✓ | 1 | |||||||||
| 89. | FUS | ✓ | ✓ | 2 | ||||||||
| 90. | G6PD | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 6 | ||||
| 91. | ASPM | ✓ | ✓ | 2 | ||||||||
| 92. | IS1 | ✓ | 1 | |||||||||
| 93. | COPD | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 94. | ERCC4 | ✓ | ✓ | 2 | ||||||||
| 95. | ERCC3 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 96. | PPARGC1A | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 97. | TUSC2 | ✓ | 1 | |||||||||
| 98. | H3P10 | ✓ | 1 | |||||||||
| 99. | ADCYAP1 | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 100. | KCNH8 | ✓ | 1 | |||||||||
| 101. | CYLD | ✓ | ✓ | ✓ | 3 | |||||||
| 102. | DPP4 | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 103. | AGTR2 | ✓ | ✓ | ✓ | 3 | |||||||
| 104. | EIF4G2 | ✓ | ✓ | ✓ | 3 | |||||||
| 105. | TMEM201 | ✓ | 1 | |||||||||
| 106. | ELK1 | 0 | ||||||||||
| 107. | EPHB2 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 108. | SENP6 | 0 | ||||||||||
| 109. | GLO1 | ✓ | ✓ | ✓ | 3 | |||||||
| 110. | ASPA | ✓ | 1 | |||||||||
| 111. | MUC1 | ✓ | ✓ | ✓ | 3 | |||||||
| 112. | MYOD1 | ✓ | 1 | |||||||||
| 113. | NFE2L2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 114. | SERPINE1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 115. | GEMIN4 | ✓ | ✓ | 2 | ||||||||
| 116. | FOXP3 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 117. | SIRT6 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 118. | PIN1 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 119. | POLG | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 120. | APTX | 0 | ||||||||||
| 121. | ENOSF1 | 0 | ||||||||||
| 122. | ASIP | ✓ | ✓ | ✓ | 3 | |||||||
| 123. | MMP9 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 9 | |
| 124. | GUSB | ✓ | 1 | |||||||||
| 125. | A1CF | ✓ | 1 | |||||||||
| 126. | H1-4 | 0 | ||||||||||
| 127. | APOE | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 128. | IGFBP3 | ✓ | ✓ | ✓ | 3 | |||||||
| 129. | IL1A | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 8 | ||
| 130. | IL1B | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 10 |
| 131. | AR | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 7 | |||
| 132. | LBR | ✓ | 1 | |||||||||
| 133. | LPA | ✓ | ✓ | ✓ | ✓ | ✓ | 5 | |||||
| 134. | MDM2 | ✓ | ✓ | ✓ | ✓ | 4 | ||||||
| 135. | MECP2 | ✓ | ✓ | 2 | ||||||||
| 136. | NHP2 | ✓ | ✓ | 2 | ||||||||
| Total number of similar genes out of 136 PMA genes | 56 | 71 | 54 | 86 | 23 | 15 | 53 | 35 | 44 | 26 |
The genes were selected from DisGeNET by entering the name of the disease (i.e., premature aging). Then, we used the DisGeNET Cytoscape App, which is an application designed for visualizing, querying, and analyzing DisGeNET data. Through a variety of built-in features, the application helps users understand genetics and explore complex human diseases and enables easy generation of disease-gene, disease-variant networks, disease-disease, and gene-gene networks. Nodes are connected if they share a neighbor in the original gene-disease network. These networks can be built around specific genes and diseases or based on the source or disease class12. For the gene network analysis of the 136 genes associated with premature aging, we found 133 nodes and 822 edges. The average node degree was 12.4, and the average local clustering coefficient was 0.512 based on the results of the STRING online network analysis tool and Cytoscape software (Figure 2 and Figure 3, respectively)13, 14.


Klotho (KL)
In early 1997, Kuro-o et al. reported that mutations in the KL gene in mice led to a syndrome that resembled aging. Mice with the aging phenotype showed growth retardation, short life spans, and atrophy of the testes, uterus, ovaries, and thymus. In the aorta and middle-sized muscular arteries, medial calcification and intimal thickening were observed. The small renal arteries also showed extensive calcification and arteriosclerosis, and ectopic calcification was found in various organs, such as the arterial walls of the stomach, bronchial mucosa, alveolar cells, choroid plexuses, skin, testes, and cardiac muscle. There was a decrease in bone mineral density and the appearance of sparse hair, emphysema, and abnormalities in growth hormone (GH), luteinizing hormone, and follicle-stimulating hormone-producing cells in the pituitary gland. Increased calcium and phosphorus levels were observed, and hypoglycaemic conditions developed due to decreased pancreatic insulin levels. Further, PBMC analysis showed that the lymphocyte-to-leukocyte ratio decreased while total protein, albumin, cholesterol, and triglyceride levels were normal. In control mice, the KL gene was highly expressed in the brain, pituitary glands, skeletal muscle, kidney, urinary bladder, pancreas, testis, and ovaries and slightly expressed in the placenta, aorta, colon, and thyroid gland. The KL gene was downregulated in the brain and kidney of mice with mutated KL genes10. Wang et al. reported that KL gene transfer to rat aorta smooth muscle (RASM) cells attenuated superoxide production and oxidative stress by suppressing Nox2 NADPH oxidase protein expression. This decreased angiotensin II(AngII)-induced superoxide production, oxidative damage, and circumvented apoptosis. However, intracellular cAMP levels and PKA activity increased in a dose-dependent manner in RASM cells15.
Werner syndrome gene (WRN)
WRN gene, also known as Werner syndrome RecQ-like helicase (WRN/RECQL2), plays major roles in genomic stability maintenance, DNA repair, replication and transcription, and telomere maintenance. Defects in this gene cause Werner syndrome, which is characterized by faster aging and higher rates of cancers (https://www.ncbi.nlm.nih.gov/gene/7486#gene-expression). In a recent study, Zimmer et al. reported that mutations in the WRN gene were found in 80 cancer primary tumor samples out of 6854 samples (1.2 %), with mutations being more prevalent in right-sided tumors. Tumor mutational burden (TMB), programmed death-ligand 1(PD-L1), and microsatellite instability-high/mismatch repair system deficient (MSI-H/dMMR) biomarkers were higher in the WRN-mutated group than the WRN-wild type group. TP53, KRAS, and APC were found in 47 (71%), 34 (49%), and 56 (73%) samples, respectively, in the WRN-mutated and WRN wild-type groups16.
Excision repair cross-complementation group 6 (ERCC6)
ERCC6 is also referred to as ERCC excision repair 6, chromatin remodeling factor/Cockayne syndrome B protein, and has been reported to be involved in DNA repair mechanisms via the nucleotide excision repair (NER) pathway. It has been reported to correct a NER defect in UV-sensitive Chinese hamster ovary (CHO) mutants. The same group also investigated Cockayne syndrome using Cellosaurus cell lines (CS1AN) with mutations in the ERCC6I gene17, 18. The ERCC6 gene was transfected to the mutants, and more UV-surviving colonies were observed than the heterogeneous colonies found in CS1AN. This indicated that ERCC6 correction occurred in the mutant cells.
Amyloid-beta precursor protein (APP)
APP is located on chromosome 21q21 and encodes a ubiquitously expressed type 1 transmembrane protein. APP generally undergoes non-amyloidogenic processing by consecutive cleavage through alpha and gamma secretases within the Aβ domain. This leads to non-pathogenic fragments, sAPP- fragments, and C-terminal fragments (CTFs). APP can be subjected to sequential proteolytic cleavage by beta and gamma secretases, which results in neurotoxic Aβ peptides (sAPP- and CTF)19. Muche et al. reported that oxidative stress increases the upregulation of APP and sAPP- β, induces VEGF synthesis, and nitric oxide oxygen free radical production. It also induces a variety of changes in endothelial phospo-p42/44 MAPK expression, which indicates that oxidative stress might play a major role in the regulation of the beta site APP cleavage enzyme-1 (BACE)20.
Exostosin glycosyltransferase 1 (EXT1)
EXT1 encodes an endoplasmic reticulum-resident type II transmembrane glycosyltransferase known to be involved in the chain elongation step of heparan sulfate biosynthesis (https://www.ncbi.nlm.nih.gov/gene/2131). EXT is genetically heterogeneous. EXT1 is located on chromosome 8q23-q24, EXT2 on chromosome 11p11-p12, and EXT3 on the short arm of chromosome 19. EXT1 is highly expressed, encodes a 746 amino acid residue protein, and possesses a tumor suppressor function. Reports have indicated that EXT1 mutations are present in chondrosarcoma21.
Lamin A (LMNA)
The LMNA gene encodes lamin A, lamin C, lamin C2, and lamin A delta 10 and is expressed in most tissues. Reports indicate that mutations in lamina/nuclear envelope proteins cause genetic diseases, such as laminopathies. Researchers have identified over 400 mutations in LMNA are responsible for diseases22. Lamin expression is reported to be regulated by the tumor suppressor p53 and retinoblastoma protein (pRb), telomere functions, regulators of the cell cycle, apoptosis, replicative senescence, and autophagy. Reports also suggest that mutant progerin lamin A, which causes Hutchinson-Gilford Progeria Syndrome (HGPS), is upregulated during normal aging23, 24. Sieprath et al. reported that the oxidation of conserved cysteine residues in the tail domain of lamin A by oxidative stress appears to impact its function and promote cellular senescence and susceptibility to reactive oxygen species (ROS). Furthermore, increased telomere shortening has been observed in most human fibroblast cell lines expressing various mutant LMNA variants. Previous studies indicate that upregulation of wild-type LMNA also causes increased telomere shortening. In laminopathies, ROS are linked to DNA damage25.
RecQ-like helicase 3 (RECQL3/BLM)
BLM or RECQL3 is an ATP-dependent RECQ DNA helicase that plays a pivotal role in the regulation of DNA replication, recombination, and homologous and non-homologous double-stranded break repair (DSBR). Mutations in BLM cause an autosomal recessive disorder known as Bloom syndrome (BS) that is characterized by genomic instability, premature aging, increased susceptibility to cancer, and immunodeficiency. BLM is located on chromosome 15q26.1 and encodes a 1417 amino acid protein-coding gene. BLM levels are higher during the S and G2 phases of the cell cycle26. Subramanian et al. reported that BLM-deficient cells collected from BS patients exhibited higher mitochondrial mass, increased mitochondrial transcription factor A (TFAM) levels, higher ATP levels, and increased respiratory reverse capacity. Increased levels of ROS and DNA base damage and an oxidative stress-dependent DNA replication rate reduction were also reported27.
RecQ like helicase 4 (RECQL4)
RECQL4 is a DNA helicase that belongs to the RecQ helicase family, which consists of other helicases, such as RECQL2 (WRN) and RECQL3 (BLM). RecQ helicases play major roles in DNA unwinding, replication, transcription, and repair. Reports indicate that mutations in these genes lead to BS, Werner syndrome, and Rothmund-Thomson syndrome (RTS) and are associated with premature aging and increased cancer prevalence28, 29. A study by Werner et al. on fibroblasts collected from RTS patients found that in normal human fibroblasts, RECQL4 was localized to the cytoplasm, with nuclear translocation and foci formation occurring in response to oxidant stimulation, while RTS fibroblasts exhibited irreversible growth arrest. RTS cells that are exposed to oxidative stress demonstrated decreased DNA synthesis, indicating that RECQL4-deficient fibroblasts were more sensitive to oxidants30.
Tumor protein p53 (TP53)
TP53 acts as a tumor suppressor and encodes the p53 transcription factor, a prominent tumor suppressor and regulator of different signaling pathways. P53 plays important roles in cell cycle arrest, DNA repair, cellular senescence, and apoptosis. TP53 mutations have been detected in most cancer types at varying prevalences, such as in 10% of hematopoietic malignancies to around 100% of high-grade serous ovarian carcinomas31.
Sirtuin 1 (SIRT1)
SIRT1 is a mammalian nicotinamide adenine dinucleotide (NAD+)0dependent histone deacetylase that acts as an epigenetic mediator of longevity, regulates DNA stability, and protects cells from oxidative stress. SIRT1 has been reported to be expressed in the brain, liver, heart, spleen, kidney, skeletal muscle, endothelial tissue, and pancreas. The expression and activation of SIRT1 leads to the modulation of its downstream pathways via targeting of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), coactivator peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), peroxisome proliferators-activated receptor-gamma (PPAR-γ), protein tyrosine phosphatase (PTP), forkhead transcriptional factors (FoxO subgroup), adenosine monophosphate-activated protein kinase (AMPK), CRE-binding protein regulated transcription coactivator 2 (CRTC2), endothelial nitric oxide synthase (eNOS), p53, myogenic differentiation (MyoD), liver X receptor (LXR), and transcription factor E2F132. Liver SIRT1 levels decrease with age, which is associated with simultaneously increased DNA damage. A similar trend has been reported in the arteries, indicating SIRT1’s association with cardiovascular diseases33.
Pathway analysis
We conducted the pathway analysis of the identified genes using the KEGG software. The genes entered were KL, WRN, ERCC6, APP, EXT1, LMNA, RECQL3/BLM, RECQL4, TP53, and SIRT1. We obtained pathways for 8 of the genes34, 35, 36. The entry identities of these genes are given below.
The results obtained from KEGG database for the different ageing or ageing related disease pathway for these genes are given in Supplement.
Discussion
Aging is related to the type of oxidative stress an individual is exposed to. Oxidative stress plays an important role in the early aging phenotype, and low-dose chronic exposure to oxidants plays an important role in many diseases, particularly cancer. In vitro studies indicate that when cells are exposed to stress conditions, they either attenuate the stress or adapt to the stress condition. Ghosh et al. reported that NADH dehydrogenase subunits are overexpressed following chronic exposure to the oxidant H2O2 37. Chronic low-dose exposure to H2O2 enhances the activity of antioxidant enzymes, such as catalase and superoxide dismutase38. The same group investigated -radiation-induced damage and found that prolonged treatment enhanced HPRT mutations in V79 cells compared to parental cells. Furthermore, conditioned cells also showed higher numbers of radiation-induced mutations39, 40. Oxidative stress has been associated with cellular senescence and colon cancer41. When cellular stress exceeds cell limits, it leads to cellular biomolecule damage. The use of antioxidants can neutralize cellular oxidants and alleviate various cellular stress-associated diseases42. Naturally occurring molecules that have antioxidant properties, such as vitamin C, vitamin E, and polyphenols (e.g., flavonols and flavanones), can be used as antioxidants to alleviate oxidative stress and neutralize oxidants43. Nanotechnology is an advanced technology that can increase the efficiency of drug delivery and drug bioavailability. Agraharam et al. reported that nanoencapsulation of a flavonol (Myricetin) led to increased bioavailability and enhanced antioxidant and antioxidant enzyme properties, such as catalase, glutathione peroxidase, and superoxide dismutase44.
To prevent replicative senescence or genomic instability, various factors are located at telomeres to regulate their length, structure, and function. There are several rare heterogeneous premature-aging diseases caused by Mendelian defects in these factors, which are known as short-telomere syndromes, telomeropathies, or telomere biology disorders (TBDs). The main functions associated with TBD-causing genes identified have been described by researchers. Studies have explored how TBDs are influenced by molecular mechanisms, including genetic anticipations, phenocopies, incomplete penetrance, and somatic genetic rescues. Studies have also discussed the diagnostic, therapeutic, and clinical implications of TBD phenotypic and genetic features, as well as the biological aspects of human telomeres, aging, and cancer45. The process of aging is complex and multifaceted. It results in the widespread functional decline of all organs and tissues; however, it is unclear whether aging is a result of a unifying cause or multiple factors. A wide range of molecular, cellular, and physiological characteristics are associated with the aging process phenotypically, such as genomic and epigenomic changes, proteostasis loss, deterioration of cell and subcellular functions, and deregulation of signaling pathways. The range of characteristics associated with this have resulted in a lack of clarity regarding their relative importance, mechanistic interrelationships, and hierarchical order. In recent years, researchers have accumulated evidence suggesting that DNA damage is responsible for most, if not all, aspects of aging. A unified approach to counteracting age-related dysfunction and disease may be achieved by improving the current understanding of DNA damage and its mechanistic relationships with aging46. There has been much discussion and improved understanding regarding how epigenetic changes occur during the aging process and how epigenetic mechanisms impact health and lifespan extension. The paper outlines questions for future research on interventions designed to rejuvenate the epigenome and delay the aging process47. Some natural compounds have been found to attenuate the aging process. Xylopia aromatica, Xylopia sericea, and other organisms contain a natural substance called myrtenal, which possesses oxidative stress-scavenging properties. In diabetic rats, myrtenal has reduced oxidative stress48.
In the present study, we identified 136 genes that are associated with different age-related diseases. Of the 136 genes, 9 had high scores in relation to their association with premature aging. This review explained the roles of these genes in aging and aging-related diseases. The present work is an in silico study. Therefore, experiments related to the expression of these genes should be undertaken and validated using various models of aging.
Conclusion
The premature aging genes that were retrieved from DisGeNET played important roles in maintaining cellular integrity, DNA repair, cell cycle regulation, cellular stress regulation, and tumor suppression pathways. The presented data indicate that mutations or age-related damage to these genes at specific sites results in age-related diseases, such as cardiovascular disease, diabetes, cancer, osteoarthritis, and osteoporosis. The data also suggests that age-related diseases are associated with aging-associated genes, and earlier genetic diagnosis may enable early treatment.
Abbreviations
AD: Alzheimer’s diseaseAMPK: adenosine monophosphate activated protein kinaseAngII: angiotensin IIARMD: Age-related macular degenerationAβ peptides: Amyloid betaBS: Bloom syndromecAMP: Cyclic Adenosine monophosphateCHO: Chinese hamster ovaryCRC: Colorectal cancerCRTC2: CRE-binding protein regulated transcription coactivator 2CS1AN: Cellosaurus cell linesCTFs: C-terminal fragmentsCVD: Cardiovascular diseasedMMR: mismatch repair system deficientDNA: Deoxyribonucleic acidDSBR: double-strand break repair eNOS: endothelial nitric oxide synthaseETC: Electron transport chain GH: Growth hormone H2O2: Hydrogen peroxideHGPS: Hutchinson-Gilford Progeria Syndrome LXR: liver X receptor MAPK: Mitogen-activated protein kinaseMSI-H: microsatellite instability-highmtDNA: Mitochondrial DNAMyoD: myogenic differentiation NADH: Nicotinamide adenine dinucleotide (NAD) + hydrogen (H)NER: Nucleotide Excision RepairNF-κB: nuclear factor kappa BetaOA: OsteoarthritisOP: OsteoporosisPBMC: Peripheral blood mononuclear cellPD: Parkinson’s diseasePD-L1: programmed death-ligand 1PGC-1α: Coactivator peroxisome proliferator-activated receptor gamma coactivator 1-alphaPKA: Protein kinase APMA: Pre-mature ageing genesPPAR-γ: Peroxisome proliferators-activated receptor-gammaPTP: Protein tyrosine phosphatase RASM: rat aorta smooth muscleRNA: Ribonucleic acidROS: reactive oxygen speciesr-Proteins: Ribosomal ProteinsrRNA: Ribosomal RNARTS: Rothmund-Thomson syndromes sAPP-∝: soluble Amyloid precursor protein - ∝sAPP-β: soluble Amyloid precursor protein - βSNHL: Sensory neural hearing lossTFAM: Mitochondrial transcription factor A TMB: Tumor mutational burdenV79 cells: Chinese hamster cellsVEGF: Vascular endothelial growth factor
Acknowledgments
We acknowledge Chettinad Academy of Research and Education for providing the CARE Seed Grant (No. Grant Number-Ref No. 004/Regr./AR-Research/2022-06).
Author’s contributions
All the authors have equally contributed to concept designing, data retrieving, analysis, literature searching, and manuscript writing. KG and AG finalized the draft. All authors read and approved the final manuscript.
Funding
Chettinad Academy of Research and Education for providing the CARE Seed Grant (No. Grant Number-Ref No. 004/Regr./AR-Research/2022-06)
Availability of data and materials
Not applicable.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
-
Harries
L.,
Goljanek-Whysall
K.,
The biology of ageing and the omics revolution. Biogerontology.
2018;
19
(6)
:
435-6
.
View Article Google Scholar -
Finkel
T.,
Holbrook
N.J.,
Oxidants, oxidative stress and the biology of ageing. Nature.
2000;
408
(6809)
:
239-47
.
View Article Google Scholar -
López-Otín
C.,
Blasco
M.A.,
Partridge
L.,
Serrano
M.,
Kroemer
G.,
The hallmarks of aging. Cell.
2013;
153
(6)
:
1194-217
.
View Article Google Scholar -
Lemoine
M.,
The evolution of the hallmarks of aging. Frontiers in Genetics.
2021;
12
:
693071
.
View Article Google Scholar -
Rattan
S.I.,
Increased molecular damage and heterogeneity as the basis of aging. Biological Chemistry.
2008;
389
(3)
:
267-272
.
View Article Google Scholar -
Cooke
M.S.,
Evans
M.D.,
Dizdaroglu
M.,
Lunec
J.,
Oxidative DNA damage: mechanisms, mutation, and disease. The FASEB Journal.
2003;
17
(10)
:
1195-214
.
View Article Google Scholar -
Jang
J.Y.,
Blum
A.,
Liu
J.,
Finkel
T.,
The role of mitochondria in aging. The Journal of Clinical Investigation.
2018;
128
(9)
:
3662-70
.
View Article Google Scholar -
Perera
R.M.,
Zoncu
R.,
The lysosome as a regulatory hub. Annual Review of Cell and Developmental Biology.
2016;
32
(1)
:
223-53
.
View Article Google Scholar -
Shcherbik
N.,
Pestov
D.G.,
The impact of oxidative stress on ribosomes: from injury to regulation. Cells.
2019;
8
(11)
:
1379
.
View Article Google Scholar -
Kuro-o
M.,
Matsumura
Y.,
Aizawa
H.,
Kawaguchi
H.,
Suga
T.,
Utsugi
T.,
Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature.
1997;
390
(6655)
:
45-51
.
View Article Google Scholar -
Sriramulu
S.,
Malayaperumal
S.,
Nandy
S.K.,
Banerjee
A.,
Essa
M.M.,
Chidambaram
S.,
Silencing of Astrocyte Elevated Gene-1 (AEG-1) inhibits the proliferative and invasive potential through interaction with Exostosin-1 (EXT-1) in primary and metastatic colon cancer cells. Biocell.
2021;
45
(3)
:
563
.
View Article Google Scholar -
Piñero
J.,
Ramírez-Anguita
J.M.,
Saüch-Pitarch
J.,
Ronzano
F.,
Centeno
E.,
Sanz
F.,
The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Research.
2020;
48
:
845-55
.
-
Szklarczyk
D.,
Gable
A.L.,
Nastou
K.C.,
Lyon
D.,
Kirsch
R.,
Pyysalo
S.,
The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Research.
2021;
49
:
605-12
.
View Article Google Scholar -
Shannon
P.,
Markiel
A.,
Ozier
O.,
Baliga
N.S.,
Wang
J.T.,
Ramage
D.,
Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Research.
2003;
13
(11)
:
2498-504
.
View Article Google Scholar -
Wang
Y.,
Kuro-o
M.,
Sun
Z.,
Klotho gene delivery suppresses Nox2 expression and attenuates oxidative stress in rat aortic smooth muscle cells via the cAMP-PKA pathway. Aging Cell.
2012;
11
(3)
:
410-7
.
View Article Google Scholar -
Zimmer
K.,
Puccini
A.,
Xiu
J.,
Baca
Y.,
Spizzo
G.,
Lenz
H.J.,
WRN-mutated colorectal cancer is characterized by a distinct genetic phenotype. Cancers (Basel).
2020;
12
(5)
:
1319
.
View Article Google Scholar -
Troelstra
C.,
Gool
A. van,
Wit
J. de,
Vermeulen
W.,
Bootsma
D.,
Hoeijmakers
J.H.,
ERCC6, a member of a subfamily of putative helicases, is involved in Cockayne's syndrome and preferential repair of active genes. Cell.
1992;
71
(6)
:
939-53
.
View Article Google Scholar -
Muftuoglu
M.,
Souza-Pinto
N.C. de,
Dogan
A.,
Aamann
M.,
Stevnsner
T.,
Rybanska
I.,
Cockayne syndrome group B protein stimulates repair of formamidopyrimidines by NEIL1 DNA glycosylase. The Journal of Biological Chemistry.
2009;
284
(14)
:
9270-9
.
View Article Google Scholar -
Hooli
B.,
Tanzi
R.E.,
The Genetic Basis of Alzheimer's Disease: Findings From Genome-Wide Studies. Genomics, Circuits, and Pathways in Clinical NeuropsychiatryElsevier 2016.
Google Scholar -
Muche
A.,
Arendt
T.,
Schliebs
R.,
Oxidative stress affects processing of amyloid precursor protein in vascular endothelial cells. PLoS One.
2017;
12
(6)
:
e0178127
.
View Article Google Scholar -
Wuyts
W.,
Hul
W. Van,
Boulle
K. De,
Hendrickx
J.,
Bakker
E.,
Vanhoenacker
F.,
Mutations in the EXT1 and EXT2 genes in hereditary multiple exostoses. American Journal of Human Genetics.
1998;
62
(2)
:
346-54
.
View Article Google Scholar -
Cenni
V.,
Capanni
C.,
Mattioli
E.,
Schena
E.,
Squarzoni
S.,
Bacalini
M.G.,
Lamin A involvement in ageing processes. Ageing Research Reviews.
2020;
62
:
101073
.
View Article Google Scholar -
Shimi
T.,
Goldman
R.D.,
Nuclear lamins and oxidative stress in cell proliferation and longevity. Cancer Biology and the Nuclear Envelope: Recent Advances May Elucidate Past Paradoxes.
2014;
2014
:
415-30
.
View Article Google Scholar -
Eriksson
M.,
Brown
W.T.,
Gordon
L.B.,
Glynn
M.W.,
Singer
J.,
Scott
L.,
Recurrent de novo point mutations in lamin A cause Hutchinson-Gilford progeria syndrome. Nature.
2003;
423
(6937)
:
293-8
.
View Article Google Scholar -
Sieprath
T.,
Darwiche
R.,
Vos
W.H. De,
Lamins as mediators of oxidative stress. Biochemical and Biophysical Research Communications.
2012;
421
(4)
:
635-9
.
View Article Google Scholar -
Kaur
E.,
Agrawal
R.,
Sengupta
S.,
Functions of BLM helicase in cells: is it acting like a double-edged sword?. Frontiers in Genetics.
2021;
12
:
634789
.
View Article Google Scholar -
Subramanian
V.,
Rodemoyer
B.,
Shastri
V.,
Rasmussen
L.J.,
Desler
C.,
Schmidt
K.H.,
Bloom syndrome DNA helicase deficiency is associated with oxidative stress and mitochondrial network changes. Scientific Reports.
2021;
11
(1)
:
2157
.
View Article Google Scholar -
Fei
F.,
Harada
S.,
Wei
S.,
Siegal
G.P.,
Molecular pathology of osteosarcoma. Bone Sarcomas and Bone Metastases-From Bench to BedsideElsevier 2022.
Google Scholar -
Sun
J.,
Wang
Y.,
Xia
Y.,
Xu
Y.,
Ouyang
T.,
Li
J.,
Mutations in RECQL gene are associated with predisposition to breast cancer. PLOS Genetics.
2015;
11
(5)
:
e1005228
.
View Article Google Scholar -
Jin
W.,
Liu
H.,
Zhang
Y.,
Otta
S.K.,
Plon
S.E.,
Wang
L.L.,
Sensitivity of RECQL4-deficient fibroblasts from Rothmund-Thomson syndrome patients to genotoxic agents. Human Genetics.
2008;
123
(6)
:
643-53
.
View Article Google Scholar -
Rivlin
N.,
Brosh
R.,
Oren
M.,
Rotter
V.,
Mutations in the p53 tumor suppressor gene: important milestones at the various steps of tumorigenesis. Genes & Cancer.
2011;
2
(4)
:
466-74
.
View Article Google Scholar -
Elibol
B.,
Kilic
U.,
High levels of SIRT1 expression as a protective mechanism against disease-related conditions. Frontiers in Endocrinology (Lausanne).
2018;
9
:
614
.
View Article Google Scholar -
Grabowska
W.,
Sikora
E.,
Bielak-Zmijewska
A.,
Sirtuins, a promising target in slowing down the ageing process. Biogerontology.
2017;
18
(4)
:
447-76
.
View Article Google Scholar -
Kanehisa
M.,
Goto
S.,
KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Research.
2000;
28
(1)
:
27-30
.
View Article Google Scholar -
Kanehisa
M.,
Toward understanding the origin and evolution of cellular organisms. Protein Science.
2019;
28
(11)
:
1947-51
.
View Article Google Scholar -
Kanehisa
M.,
Furumichi
M.,
Sato
Y.,
Kawashima
M.,
Ishiguro-Watanabe
M.,
KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Research.
2023;
51
:
587-92
.
View Article Google Scholar -
Ghosh
R.,
Girigoswami
K.,
NADH dehydrogenase subunits are overexpressed in cells exposed repeatedly to H2O2. Mutation Research.
2008;
638
(1-2)
:
210-5
.
View Article Google Scholar -
Girigoswami
K. Bose,
Bhaumik
G.,
Ghosh
R.,
Induced resistance in cells exposed to repeated low doses of H2O2 involves enhanced activity of antioxidant enzymes. Cell Biology International.
2005;
29
(9)
:
761-7
.
View Article Google Scholar -
Girigoswami
K. Bose,
Ghosh
R.,
Response to γ-irradiation in V79 cells conditioned by repeated treatment with low doses of hydrogen peroxide. Radiation and Environmental Biophysics.
2005;
44
(2)
:
131-7
.
View Article Google Scholar -
Bose
K.,
Bhaumik
G.,
Ghosh
R.,
Chronic low dose exposure to hydrogen peroxide changes sensitivity of V79 cells to different damaging agents. Indian Journal of Experimental Biology.
2003;
41
(8)
:
832-6
.
-
Bhatiya
M.,
Pathak
S.,
Banerjee
A.,
Oxidative Stress and Cellular Senescence: The Key Tumor-promoting Factors in Colon Cancer and Beneficial Effects of Polyphenols in Colon Cancer Prevention. Current Cancer Therapy Reviews.
2021;
17
(4)
:
292-303
.
View Article Google Scholar -
Agraharam
G.,
Girigoswami
A.,
Girigoswami
K.,
Myricetin: a Multifunctional Flavonol in Biomedicine. Current Pharmacology Reports.
2022;
8
(1)
:
48-61
.
View Article Google Scholar -
De
S.,
Gopikrishna
A.,
Keerthana
V.,
Girigoswami
A.,
Girigoswami
K.,
An Overview of Nanoformulated Nutraceuticals and Their Therapeutic Approaches. Current Nutrition and Food Science.
2021;
17
(4)
:
392-407
.
View Article Google Scholar -
Agraharam
G.,
Girigoswami
A.,
Girigoswami
K.,
Nanoencapsulated myricetin to improve antioxidant activity and bioavailability: a study on zebrafish embryos. Chemistry (Basel, Switzerland).
2021;
4
(1)
:
1-17
.
View Article Google Scholar -
Revy
P.,
Kannengiesser
C.,
Bertuch
A.A.,
Genetics of human telomere biology disorders. Nature Reviews. Genetics.
2023;
24
(2)
:
86-108
.
View Article Google Scholar -
Schumacher
B.,
Pothof
J.,
Vijg
J.,
Hoeijmakers
J.H.,
The central role of DNA damage in the ageing process. Nature.
2021;
592
(7856)
:
695-703
.
View Article Google Scholar -
Zhang
W.,
Qu
J.,
Liu
G.H.,
Belmonte
J.C.,
The ageing epigenome and its rejuvenation. Nature Reviews. Molecular Cell Biology.
2020;
21
(3)
:
137-50
.
View Article Google Scholar -
Rathinam
A.,
Pari
L.,
Venkatesan
M.,
Munusamy
S.,
Myrtenal attenuates oxidative stress and inflammation in a rat model of streptozotocin-induced diabetes. Archives of Physiology and Biochemistry.
2022;
128
(1)
:
175-83
.
View Article Google Scholar
Comments
Downloads
Article Details
Volume & Issue : Vol 10 No 7 (2023)
Page No.: 5783-5795
Published on: 2023-08-03
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 4092 times
- PDF downloaded - 1142 times
- XML downloaded - 114 times
- Supplement downloaded - 938 times
