

Copyrights: Siti Farizan Mansor, Tengku Muhamad Faris Syafiq, Badrul Hisham Yahaya, 2024. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Despite the high number of cases reported, studies describing the pathogenesis of urothelial carcinoma (UC) remain limited. This is due to insufficient in vivo models that can accurately recapitulate the pathogenesis of UC recurrence and elucidate the involvement of the tumour microenvironment (TME) during carcinogenesis. Models of cancer pathomechanism in monolayer culture provide inaccurate resemblance due to biophysical and chemical changes. In simplified 2D culture conditions, factors such as tissue architecture, cell-to-cell and cell-to-matrix interaction, and mechanical and biochemical networks, all of which are involved in drug response, are lost. Three-dimensional (3D) culture of clinical biopsy is considered an ideal model to understand UC pathogenesis, the role of the microenvironment, and mechanical adaptations due to improved translational capacity. However, limited biopsies and challenges in primary cell culturing have shifted researchers towards developing cell line-based 3D culture in the pursuit of pathomechanism exploration. A comprehensive literature evaluation was carried out by searching the PubMed, Scopus, and Web of Science databases from January 2000 to May 2022. The initial search yielded 525 articles, and 195 studies were selected based on the inclusion and exclusion criteria. This review highlights recent challenges, future strategies, and the clinical implications of developing high-throughput, cell line-based 3D models for personalized UC treatment, a critical gap in the current literature.
Introduction
Urothelial carcinoma (UC) is the second most frequent urogenital cancer after prostate cancer, which among all cancers, was ranked eleventh globally1. Almost 30% of UC patients develop recurrence within two years of treatment2. This high recurrence rate makes UC one of the most expensive cancers to manage. The treatment costs reach up to US$162,574 per patient annually in the US and Canada, surpassing breast (US$85,772) and lung cancers (US$135,567)3, 4, 5.
Understanding UC recurrence is hindered by inadequate in vivo models and the inability to effectively study the tumor microenvironment. Traditional 2D monolayer culture fails to accurately model cancer etiology, as they lack cell-to-cell and cell-to-matrix interactions, tissue architecture, metabolic networks, and mechanical properties. A 3D model has proven critical not only for UC but also for advancing understanding in various cancers such as prostate and breast cancer6. Moreover, it provides enhanced accuracy in mimicking the TME, understanding metastasis, and improving research outcomes in drug testing. Cells can sense dimensionality and other physical and biological features of the ECM via integrins. Cells react differently to 2D and 3D substrates, triggering various signaling pathways in each7. Protease activation which leads to extracellular matrix (ECM) degradation is hindered in 2D models8. Conversely, cells in 3D cultures can sense and respond to ECM features, triggering distinct signaling pathways compared to 2D cultures7. For instance, 3D cultures show higher expression of integrins and MMP9, which are crucial for cell spheroid formation9. Although animal models offer advantages over 2D cultures, issues of reproducibility, scalability, and cost limit their use10.
3D culture models provide a more accurate representation for studying UC pathogenesis by maintaining the tumor microenvironment (TME) and multicellularity. They allow genetic and phenotypic analysis of sequential events during pathogenesis, thus clarifying the disease cascade. Moreover, 3D cultures could better facilitate research on cancer cell proliferation, migration, invasion, and response to chemo- and radio-therapy. Co-culturing spheroids with components of the TME, such as stromal and immune cells, helps in understanding how tumors adapt to their microenvironment. For example, Martinelli (2022) showed that SDHB and SDHD silenced pheochromocytoma spheroids exhibit different behaviors when interacting with the TME, with impaired stroma metabolism inhibiting tumor aggressiveness11. Spheroid models allow researchers to explore tumor-TME interactions and discover potential therapeutic targets, which are crucial for developing effective cancer treatments.
In addition to radiotherapy, other drugs, such as metformin, can significantly affect spheroid patterns and interactions within the TME. Martinelli (2022) demonstrated that metformin induces distinct responses in pheochromocytoma/paraganglioma tumor cells and primary fibroblasts. The study showed that metformin differentially impacted tumor spheroids and fibroblasts within the TME, altering their metabolic profiles and reducing tumor aggressiveness. Specifically, metformin suppressed proliferation in tumor cells while inducing changes in fibroblast metabolism, demonstrating how drugs can modulate both tumor cells and TME components12. This demonstrated that drugs not only directly target cancer cells but also alter TME dynamics, which can influence overall tumor behavior. This highlights the importance of considering drug effects on multiple cell types within the TME when evaluating treatment responses.
3D cell cultures are categorized into spheroid and organoid models. Spheroids arise from either single-cell proliferation by a monotype of cell or through multi-cell aggregation by multiple types of cells, the latter resulting from natural cell tendencies to agglomerate in suspension cultures13, 14, 15. Self-renewal in spheroids can be demonstrated by culturing cells at very low densities in ultra-low attachment microwells, allowing the identification of cells with self-renewal capabilities16. Organoid culture is a cell culture method to recapitulate 3D architecture resembling an organ. Unlike organotypic culture which involves re-culturing a whole organ derived from an organism, organoid culture is grown from multicellular and conducive biochemical factors to develop an organ-like structure17, 18. An organoid may exhibit three characteristics: self-organization, multicellularity, and functionality19. Firstly, stem cells of the organ of interest are cultured in culture media with optimal supplements to promote growth and differentiation. Within 14-30 days, the culture will grow and arrange itself in a 3D structure resembling an organ which demonstrates partial or complete organ function17.
Various methods have been developed to grow organoids for research and therapeutic purposes. Lung organoids from induced pluripotent stem cells (iPSCs) are used to study COPD and lung transplants20, 21. Neurological disorders are studied using brain organoids that are developed through the differentiation of iPSCs into organoids. The organoid demonstrates various brain cell types and specific brain regions capable of replicating physiological and pathological conditions of the blood-brain barrier22. Small intestine organoids are used for disease modeling, regenerative medicine, drug screening, and gene repair studies23.
The human urinary bladder, lined by a specialized stratified epithelium, comprises three layers: the luminal layer with umbrella cells, the intermediate layer with polygonal cells, and the basal layer with cuboidal cells and bladder stem cells. This is followed by connective tissues, muscle, and fat layers. This structure reorganizes from 5-7 layers when relaxed to 2-3 layers when the bladder is full without structural damage, illustrating its transitional epithelium nature24.
Although organoid development of various human organs has been recognized since early 2015, the organoid culture of the urinary bladder is still lacking. Bladder organoid cultures must replicate this tissue organization, forming 4-6 closely packed cell layers with distinct basal, intermediate, and luminal layers capable of retaining urine and preventing its permeabilization into the underlying structure25. Self-renewal and differentiation are the primary characteristics of human adult stem cells that underpin organoid culture26, 27. By seeding stem cells on epithelial stroma in ideal conditions, researchers can expand and differentiate these cells using genetic tools, producing tissue-like structures for research and therapeutic applications.
Spheroids are commonly derived from differentiated tumor cell lines and do not require stem cells for their formation. In contrast, organoids rely on stem cells for their long-term maintenance in vitro, as they retain high replicative potential. Stem cells and cancer stem cells (CSCs) are subpopulations found in tissue biopsies. Due to limited biopsy samples, researchers often use cell lines for 3D culture development. Non-cancerous cell lines lack or have limited stem cell populations compared to cancerous cell lines, which possess CSC subpopulations capable of developing into spheroids and organoids28.
An accurate and cost-effective model to study UC biology and chemotherapy resistance mechanisms is urgently needed, as these are major causes of recurrence29. The UC organoid culture model holds potential for evaluating existing and novel therapeutic targets, thus personalizing UC management and enhancing convenience for patients and healthcare providers. This review reports current advances in the 3D culture of UC cell lines, highlighting its applications, limitations, challenges, and future perspectives.
Methods
Article Search and Selection Strategies: The electronic databases PubMed, Scopus, and Web of Science were searched using the keywords “3D culture,” “urothelial carcinoma cell line,” and “spheroid.” A complete list of the search strings used is presented in the supplementary file. The search was restricted to articles published from January 2000 to April 2022 and in English. The articles were screened for inclusion based on the following exclusion criteria: focus on 3D brachytherapy, 3D ultrasound, 3D radiotherapy, 3D PET and CT images, immunohistochemical 3D reconstruction, 3D modeling from anatomical images, non-UC cell lines, non-original research or review articles, non-English articles, and those without available full texts. Figure 1 depicts an outline of the literature search in a PRISMA flow diagram.




Results
The search produced 525 hits, of which 473 articles were deemed eligible based on the abstract, and 195 were included after reference check and full-text screening. The publication year ranged from January 2000 to April 2022. Based on the analysis of 195 studies, several key trends were observed regarding the approaches used to develop 3D UC culture models (supplementary data). Early research mostly used simpler approaches such as hanging drop (HD), methylcellulose (MC), and collagen-based scaffolds. However, there has been a recent trend towards the use of more advanced materials and techniques. Complex platforms such as microfluidic chips and ultra-low attachment dishes (ULA) are more favored because they provide a more accurate simulation of the TME.
Besides, there has been a growing trend towards multimodal approaches. Researchers are increasingly combining various methods to enhance the accuracy of 3D models. For instance, combining ULA with Matrigel (ULA+M) or collagen with fibronectin (ULA+coll+fibronectin) demonstrates an effort to mimic both the physical and biochemical properties of the TME more comprehensively. Additionally, there has been a clear shift towards scaffold-free culturing techniques, particularly the use of ULA and poly 2-hydroxyethyl methacrylate (Poly HEMA), which minimize the potential interference with the cellular behavior caused by scaffolds.
There is also advancement in integrating biomimetic designs and microfluidic technology into 3D UC models. This innovation improves the physiological relevance of the models by demonstrating dynamic conditions, such as nutrient flow and waste elimination, better mimicking the in vivo TME. Another significant development has been the increasing use of coculture systems, where UC cells are co-cultured with other cell types, such as fibroblasts and human umbilical vein endothelial cells (HUVECs). This approach helps replicate cellular interactions within the TME and provides valuable insights into cancer progression and metastasis.
Discussion
3D culture approaches in UC cell lines
To date, there are hundreds of approaches and methods to culture UC cell lines under 3D conditions for various applications (Supplementary Tables 1 and 2). The earliest literature described the forced floating method, where cells are cultured in suspensions, independent of surface attachments30. There are three ways to suspend cells: by culturing in ULA wells, poly-hydroxyethyl methacrylate (poly-HEMA) pre-coated dishes and the hanging drop (HD) method. In the HD method, 10-20µL cell suspensions were placed on a plate. The plate was then flipped upside down, which pulled down cells to spontaneously form aggregates at the bottom of the drop13, 31. This method is also used to generate embryoid bodies from embryonic stem cells. Alternatively, the rotary culturing system enables spontaneous cell aggregation32. Another approach, static liquid overlay, induces cells to aggregate instead of sticking to a surface when suspended cells are cultivated on a nonadherent substrate33. Spheroids can also be formed by the air-liquid interface (ALI) system, where the bottom and upper parts are separately exposed to liquid and air, respectively34. Some researchers combined forced floating and liquid overlay techniques to obtain the desired outcome based on their research objectives. More recently, spheroids and organoids have been grown on nonadherent microfluidic chips35.
Limitations of 3D bladder organoid culture methods
Although organoid culture of various organs was introduced in 2000, there was limited research focused on developing a complete structural organization and functional urinary bladder organoid using cell lines. Among the 195 selected articles, a majority discussed the spheroid culture method, which we considered a very well-established method. However, less than 10 papers discussed the organoid culture method. They defined urinary bladder organoid as 4-6 layers of stratified urothelium on the surface of 2-3 mm3 of either collagen, de-epithelialized stroma, Matrigel™ or reconstituted basement membrane, with the presence of tight and gap junctions on the surface of the cells31, 36, 37, 38. The most sophisticated bladder organoid was a 3D human vesical equivalent of the bladder which was created from primary cells. Histological investigation revealed morphological similarities to natural human mucosa with the presence of ECM, a capillary-like network, basement membrane and tight junctions in the mature stratified urothelium. The vesical equivalent was then transplanted with T24 and RT4 cell lines to study their invasive potential31. This review reported that there are limited studies performed to demonstrate organoid differentiation into a stratified urothelium layer, which is 9/195 (4.6%)28, 34, 36, 39, 40, 41, 42, 43, 44 with no reports describing methods to develop functional urinary bladder vesical structure. Novel analysis and validation methods for assessing urine storage cavity during contraction will facilitate its translational potential.
Future perspectives of 3D bladder organoid culture
3D culture application in cancer chemoresistance study
Due to the attribution of UC as a stem cell disease, the primary clinical challenges in UC include identifying early diagnostic markers, prognostic indicators, and novel therapeutic targets. Investigations into the molecular signatures of UC stem cells have revealed significant intrinsic plasticity and heterogeneity, which greatly affect individual responses to therapy. Incorrect cytotoxic treatments can lead to the enrichment of cancer stem cells (CSCs), contributing to increased chemoresistance, a major obstacle in UC treatment. Therefore, a thorough understanding of the fundamental processes underlying UC stemness and their link to drug resistance is essential. As treatment approaches for UC evolve, there is a favorable shift toward more personalized therapies. Identifying unique early diagnostic and prognostic CSC markers in UC may offer new methods for disease classification or clinical staging, with the presence of CSCs potentially indicating a more aggressive cancer phenotype. Patient-derived tissue, organoid, and xenograft models of bladder cancer, representing various stages and histological subtypes, have been developed45, 46, 47, 48. These models provide insights into oncogene mutations and expression patterns unique to each patient, facilitating personalized therapy. In 2018, 16 bladder cancer organoid lines were created from patients with a 70% success rate, accurately replicating the genetic and histological properties of the original tumors45. Another study produced 133 organoids from 53 bladder cancer patients with around 50% efficiency46. These studies linked drug resistance to tumor growth, observed varied drug responses among organoid lines45, 46, and noted increased drug resistance in 3D compared to 2D cultures48. Notably, the mutational profiles of individual organoids helped identify therapeutic vulnerabilities, predicting medication responses45, 46.
3D culture application in cancer immunotherapy
Co-culture of patient bladder cancer organoids with chimeric antigen receptor T cells (CAR-T) is effective in immunotherapy evaluation49. In a co-culture system, CAR-T cells designed to target MUC1 in organoids caused death in MUC1-expressing organoids49. It is exciting that organoid models show potential as a therapeutic decision-making tool, albeit further advancement will benefit patient outcomes. Many factors influence organoid production and long-term passaging success rates, including organoid culture conditions (e.g., presence of serum and growth hormones, medium composition, kinds and sources of matrix proteins)45, 46, tumor grade/stage50, and stem cell marker expression51. These variables can have a varied impact on the growth and stability of each organoid model. Furthermore, co-culture methods containing immune cells and stromal compartments can more effectively mimic a physiologically relevant tumor microenvironment52, 53. As a result, an ideal organoid model with functional vesical structure may improve the spatiotemporal study of intravesical-based therapeutic modalities. Organoid model analyses show potential as a guide for clinical decisions and improving patient outcomes. Novel CSC-targeted treatments are still required to improve UC therapeutic efficacy. The success of UC therapy and clinical care is mainly reliant on disease models. The ideal platform for developing and establishing an efficient and accurate organoid model system is through utilization of UC cell lines. Once the system is well-established, patient-derived UC organoids could constitute a reliable model system for investigating tumor evolution and treatment response in the context of personalized cancer treatments.
3D culture feasibility in clinical settings
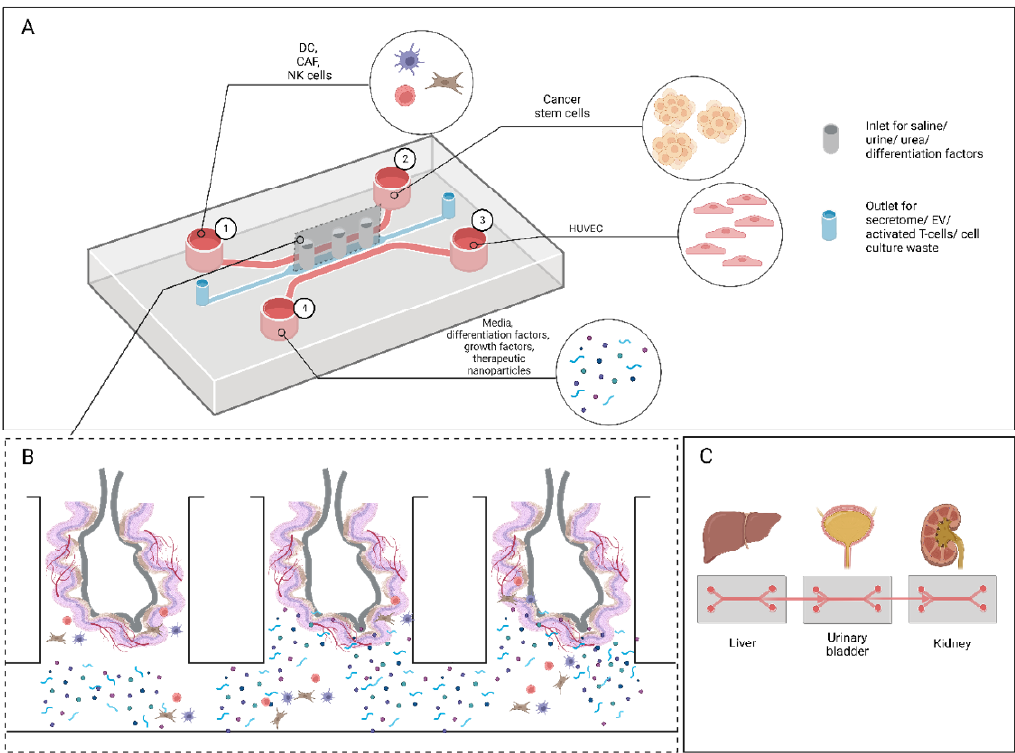
Ideally, an organoid system for studying UC should be able to demonstrate metastatic cascade stages, angiogenesis, a fully operational immune system, multicellular cell-cell and cell-ECM interactions. For the model to be cost-effective, user-friendly, and feasible to be applicable in clinical settings, it should encompass high throughput sampling with a high level of extrinsic control and self-automation which allows for conducive administration of cellular components, delivery of drugs, proteins, growth factors and differentiation factors. Additionally, the model should feature an accurate topology of the tumor components which allows precise tracking of individual cells. Besides, integration of high-resolution real-time imaging will improve the quality and acquisition of data collection (Figure 2). Interdisciplinary approaches have potential to advance the 3D culture models in UC. Combining tissue engineering with computational modeling, particularly molecular docking, allows for prediction of drug-ligand/ potential targeted protein interactions and tumor behavior. This enables the identification of promising therapeutic compounds in silico, reducing the need for exhaustive laboratory testing. Besides, the integration of machine learning provides a powerful tool for predicting patient outcomes based on vast datasets derived from 3D models. By training algorithms on patient-specific molecular profiles and treatment responses, machine learning can offer personalized therapeutic insights, therefore enhancing the clinical relevance of 3D models.
3D culture in tumor-on-a-chip devices
3D bioprinting allows for the precise deposition of cells, bioactive compounds, biomaterials, and other cellular components to create tissue-like structures that mimic in vivo. It is capable of recreating the intrinsic architecture of tissues, improving the accuracy of models used in drug testing, disease modeling, and tissue engineering. In cancer research, 3D bioprinting can produce complex TME for studying tumor behavior, drug resistance, and potential therapies that support personalized medicine54. Microfluidic advances have enabled the isolation and culture of thousands of single cells on a chip. However, the tumorsphere assay is time-consuming, limiting its throughput. Chen et al. utilized machine learning with single-cell analysis to expedite the tumorsphere assay, demonstrating potential applications in various biomedical fields55. Microfluidic systems enhance readouts by allowing high-throughput screening of therapeutic modalities in patient-derived organoids. Co-culturing with microvasculature in tumor-on-a-chip devices for in vitro chemosensitivity assays can comprehensively evaluate anticancer drug absorption, distribution, metabolism, excretion, and toxicity, closely mimicking the in vivo tumor microenvironment. Evaluating drug efficacy and safety is crucial for assessing cellular toxicities, facilitating drug screening and delivery. An ideal system should measure cellular toxicity in the liver, kidney, and blood vessels during or after drug administration in the bladder organoid. Organ-on-a-chip models emphasize hepatocyte contribution to drug metabolism by assessing adverse effects produced by drug metabolites rather than the drug itself. A prodrug is an inactive substance converted into an active form through metabolic or physico-chemical transformation to perform therapeutic actions56. For example, thymosin fraction 5 (TF5) converts to fluorouracil (5-FU), causing hepatotoxicity, while doxorubicin converts to doxorubicinol, causing hematological toxicity. Therefore, liver, kidney, and other related organs should be cultured in separate channels to examine safety and toxicity following drug administration in the bladder organoid. Co-organ systems such as liver-uterine-bone marrow57, liver-heart-vascular58, liver-intestine-breast59, liver-prostate-kidney60, liver-breast-cervical61, and liver-lung-kidney-fat62 have been utilized in organ-on-a-chip systems. These systems use homotypic hepatocarcinoma cells to study liver interactions with other organs cultured in separate chambers connected via microfluidic channels63. Tissue engineering scaffolds provide 3D environments for cell propagation and specialized signaling molecules that mimic real tissue surroundings. These scaffolds can be natural, synthetic, or a combination of both. Signaling molecules significantly enhance scaffold efficacy by controlling the phenotypes of enclosed and surrounding cells. In cellular strategies, cells are cultured in vitro and allowed to connect with scaffolds before implantation. In acellular strategies, living host cells migrate and infiltrate the implanted scaffold. These cells can then be extracted as tissues for wound healing, secondary site explants, or newly differentiated tissues from stem cell sources64. Once the scaffold is completely reabsorbed, only the newly regenerated tissue remains. The regenerated tissue allows the infiltration of angiogenesis and nerve fibers and the deposition of extracellular matrix (ECM) components. Signaling molecules within the scaffolds support and regulate innate cells such as CSCs, endothelial progenitors, and macrophages from the bloodstream and surrounding tissue64.
Elasticity is described as the capacity of a material to rebound from distortion when exposed to a force. The mechanical work required to distort the object is stored as strain energy, which can be recovered once the force is released. Resilience is the ability of a material to absorb energy after elastic deformation, with energy recovery upon unloading65. The level of resilience of a material must be measured before being used as a scaffold. This will ensure that it has adequate resilience proficiency to support the tissue as it grows into an expanding structure. Elastomers are rubber-like compounds that demonstrate elasticity. Polymer chains in these materials are linked by relatively weak intermolecular interactions thereby allowing the polymers to stretch in response to microscopic forces66. Polyglycerol sebacate (PGS) is an example of an elastomer that has been used as a scaffold for tissue engineering of cardiac tissue. They possess excellent elasticity, biocompatibility, biodegradability, and mild cytotoxicity67. Chen and colleagues have designed a heart patch from a biocompatible and biodegradable PGS. Plastering the heart patch to a damaged left ventricular wall influenced cardiac mechanics by lowering myofibril stresses and confounding changes in clinical left ventricular metrics. The reduction in wall stress was calculated to be proportional to the fractional volume added, with firmer materials improving this reduction better68, 67.
Elastomer use in bladder organoid culture was never reported. In terms of validation of an organoid model, accurate structure and function are crucial. The bladder vesical should function as a urine storage cavity that is able to expand when it is in full volume and contract when vacant. Therefore, we proposed elastomer to be used as a scaffold to support the development of bladder organoid. As the urothelial layers are growing, saline is added in increasing volume, day by day, in order to give more space for urothelial expansion. Once the organoid is fully developed surrounding the elastomer, the elastomer should be able to be naturally degraded, leaving multilayers of stratified urothelium to form a structured, organized and functional urine-filled cavity (Figure 3, Figure 4). The tight junction of umbrella cells and GAG layer account for the luminal impermeability capability of bladder organoid. To test luminal impermeability function, bladder organoid can be cultured in a filter insert as an air-liquid interface setting. The vesical can be filled up with a solution with a predetermined urea concentration and left for a few days to allow for urea to permeabilize the luminal layer, breaching the filter insert and ending up at the bottom of the culture vessel. A spectrophotometer can then be used to measure the concentration of urea at the bottom. For negative control, the luminal layer will be pretreated with acetone or SDS to disrupt tight junctions in umbrella cells. If the organoid has an intact luminal impermeability function, urea should not breach the luminal layer, thus no urea should be found at the bottom of the vessel. The Technology Readiness Levels (TRL) of the 3D culture model for urothelial carcinoma currently stand at the early to mid-range level, approximately TRL 4 to 5. Despite the promise of the proposed model in replicating tumor architecture and cell interactions in vitro, they remain in the preclinical research phase. Significant efforts are required to enhance the model’s reproducibility, scalability, and ability to mimic in vivo conditions accurately. To progress towards clinical application (TRL 6–7), further validation through large-scale testing and regulatory evaluation is necessary. This will include proving the model’s robustness and reproducibility in different laboratories, as well as demonstrating its utility in predicting patient-specific treatment outcomes. These need to be analyzed before it can be integrated into routine cancer treatment and management.
Conclusions
Development of an ideal organoid model will aid in closing the gap between simplistic 2D monolayer culture and complicated transgenic animal host. Biomimetic design inspires researchers to keep innovating and improving the design of current 3D culture models. Once the system is well-established, patient-derived UC organoids will constitute a sophisticated and accurate model system for investigating tumor evolution and treatment response in the context of personalized cancer treatments. Besides, the platform has the potential to be developed for the regeneration of bladder organs, which will greatly benefit patients with radical cystectomy.
Future research should focus on integrating interdisciplinary approaches, such as tissue engineering with computational tools. Molecular docking can be applied to study interactions of drugs and therapeutic targets, while machine learning algorithms can predict patient-specific outcomes. The incorporation of microfluidic chips will enable high-throughput screening, offering a high degree of external control and self-automation, which are keys for clinical translation. Furthermore, future research could also explore the interaction between the urothelium and endogenous microbiota. Given the potential oncogenic role of microbiota, organoid models offer a promising platform to study these interactions. Incorporating such studies may provide additional insights into tumor biology and open new avenues for therapeutic interventions.
These advancements in 3D UC models could reduce dependency on animal models, improve drug testing accuracy, and lead to more personalized and effective treatments. Robust UC 3D models promise to revolutionize therapeutic approaches and provide tangible clinical benefits for patients. The combination of emerging technologies, such as machine learning and microfluidic platforms, further enhances the potential for these models to be implemented in healthcare, pushing the boundaries of precision medicine.
Abbreviations
2D: Two-dimensional, 3D: Three-dimensional, 5-FU: Fluorouracil, ALI: Air-liquid interface, CAR-T: Chimeric antigen receptor T cells, COPD: Chronic obstructive pulmonary disease, CSCs: Cancer stem cells, ECM: Extracellular matrix, HD: Hanging drop, iPSCs: Induced pluripotent stem cells, MC: Methylcellulose, MMP9: Matrix metallopeptidase 9, MUC1: Mucin 1, PGS: Polyglycerol sebacate, Poly HEMA: Poly 2-hydroxyethyl methacrylate, PRISMA: Preferred Reporting Items for Systematic Reviews and Meta-Analyses, SDHB: Succinate dehydrogenase complex iron sulfur subunit B, SDHD: Succinate dehydrogenase complex subunit D, TF5: Thymosin fraction 5, TME: Tumour microenvironment, TRL: Technology Readiness Levels, UC: Urothelial carcinoma, ULA: Ultra-low attachment
Acknowledgments
None.
Author’s contributions
Siti Farizan Mansor: Design & Conceptualization, Investigation, Writing - Original draft, table & figure. Tengku Muhamad Faris Syafiq: Writing - Original Draft, Writing - Review & Editing. Badrul Hisham Yahaya: Writing - Review & Editing. All authors read and approved the final manuscript.
Funding
This study was funded by Ministry of Higher Education Malaysia through Fundamental Research Grant Scheme (DP KPT FRGS/1/2019/SKK15/UITM/03/1), Project ID 15531, RMC file number 600-IRMI/FRGS 5/3 (286/2019).
Availability of data and materials
Data and materials used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
-
Jemal
A.,
Bray
F.,
Center
M.M.,
Ferlay
J.,
Ward
E.,
Forman
D.,
Global cancer statistics. CA: a Cancer Journal for Clinicians.
2011;
61
(2)
:
69-90
.
View Article PubMed Google Scholar -
Feldman
A.S.,
Surgical challenges and considerations in Tri-modal therapy for muscle invasive bladder cancer. Urologic Oncology: Seminars and Original Investigations [Internet].
2021;
000
:
1-9
.
View Article PubMed Google Scholar -
Blumen
H.,
Fitch
K.,
Polkus
V.,
Comparison of treatment costs for breast cancer, by tumor stage and type of service. American Health & Drug Benefits.
2016;
9
(1)
:
23-32
.
PubMed Google Scholar -
Cheung
M.C.,
Earle
C.C.,
Rangrej
J.,
Ho
T.H.,
Liu
N.,
Barbera
L.,
Impact of aggressive management and palliative care on cancer costs in the final month of life. Cancer.
2015;
121
(18)
:
3307-15
.
View Article PubMed Google Scholar -
Santos
F.,
Dragomir
A.,
Zakaria
A.S.,
Kassouf
W.,
Aprikian
A.,
Predictors of costs associated with radical cystectomy for bladder cancer: A population-based retrospective cohort study in the province of Quebec, Canada. Journal of Surgical Oncology.
2016;
113
(2)
:
223-8
.
View Article PubMed Google Scholar -
Manduca
N.,
Maccafeo
E.,
De Maria
R.,
Sistigu
A.,
Musella
M.,
3D cancer models: one step closer to in vitro human studies. Frontiers in Immunology.
2023;
14
.
View Article PubMed Google Scholar -
Larsen
M.,
Artym
V.V.,
Green
J.A.,
Yamada
K.M.,
The matrix reorganized: extracellular matrix remodeling and integrin signaling. Current Opinion in Cell Biology.
2006;
18
(5)
:
463-71
.
View Article PubMed Google Scholar -
Mitra
A.P.,
Bartsch
J. Georg,
Cote
R.J.,
Risk Factors and Molecular Features Associated with Bladder Cancer Development. In: Hansel, D., Lerner, S. (eds) Precision Molecular Pathology of Bladder Cancer. Molecular Pathology Library. Springer, Cham. https://doi.org/10.1007/978-3-319-64769-2_1. 2017
.
-
Peng
C.C.,
Chyau
C.C.,
Wang
H.E.,
Chang
C.H.,
Chen
K.C.,
Chou
K.Y.,
Cytotoxicity of ferulic Acid on T24 cell line differentiated by different microenvironments. BioMed Research International.
2013;
2013
(1)
:
579859
.
View Article PubMed Google Scholar -
Jilka
R.L.,
The Road to Reproducibility in Animal Research. Journal of Bone and Mineral Research : the Official Journal of the American Society for Bone and Mineral Research.
2016;
31
(7)
:
1317-9
.
View Article PubMed Google Scholar -
Martinelli
S.,
Riverso
M.,
Mello
T.,
Amore
F.,
Parri
M.,
Simeone
I.,
SDHB and SDHD silenced pheochromocytoma spheroids respond differently to tumour microenvironment and their aggressiveness is inhibited by impairing stroma metabolism. Molecular and Cellular Endocrinology.
2022;
547
:
111594
.
View Article PubMed Google Scholar -
Martinelli
S.,
Amore
F.,
Mello
T.,
Mannelli
M.,
Maggi
M.,
Rapizzi
E.,
Metformin treatment induces different response in pheochromocytoma/paraganglioma tumour cells and in primary fibroblasts. Cancers (Basel).
2022;
14
(14)
:
3471
.
View Article PubMed Google Scholar -
Hairuddin
O.N.,
Yahaya
B.H.,
Ibahim
M.J.,
Verakumarasivam
A.,
Choy
C.S.,
Mazlan
M.,
Comparison of cancer stem cell enrichment between spheroids derived from single-cell and multicellular aggregate cultures. Biomedical Research and Therapy.
2023;
10
(8)
:
5810-30
.
View Article Google Scholar -
Shahbazi
M.N.,
Mechanisms of human embryo development: from cell fate to tissue shape and back. Development (Cambridge, England).
2020;
147
(14)
:
dev190629
.
View Article PubMed Google Scholar -
Martinelli
S.,
Cantini
G.,
Propato
A.P.,
Bani
D.,
Guasti
D.,
Nardini
P.,
The 3D in vitro Adrenoid cell model recapitulates the complexity of the adrenal gland. Scientific Reports.
2024;
14
(1)
:
8044
.
View Article PubMed Google Scholar -
Wang
X.,
Lin
G.,
Martins-Taylor
K.,
Zeng
H.,
Xu
R.H.,
Inhibition of caspase-mediated anoikis is critical for basic fibroblast growth factor-sustained culture of human pluripotent stem cells. The Journal of Biological Chemistry.
2009;
284
(49)
:
34054-64
.
View Article PubMed Google Scholar -
Sachs
N.,
Clevers
H.,
Organoid cultures for the analysis of cancer phenotypes. Current Opinion in Genetics & Development.
2014;
24
:
68-73
.
View Article PubMed Google Scholar -
Schweiger
P.J.,
Jensen
K.B.,
Modeling human disease using organotypic cultures. Current Opinion in Cell Biology.
2016;
43
:
22-9
.
View Article PubMed Google Scholar -
Lancaster
M.A.,
Knoblich
J.A.,
Organogenesisin a dish: Modeling development and disease using organoid technologies. Science.
2014;
345
(6194)
:
1247125
.
View Article Google Scholar -
Hsia
G.S.,
Esposito
J.,
Rocha
L.A. da,
Ramos
S.L.,
Okamoto
O.K.,
Clinical Application of Human Induced Pluripotent Stem Cell-Derived Organoids as an Alternative to Organ Transplantation. Stem Cells International.
2021;
2021
(1)
:
6632160
.
View Article PubMed Google Scholar -
Ahmed
E.,
Fieldes
M.,
Bourguignon
C.,
Mianné
J.,
Petit
A.,
Jory
M.,
Differentiation of Human Induced Pluripotent Stem Cells from Patients with Severe COPD into Functional Airway Epithelium. Cells.
2022;
11
(15)
:
2422
.
View Article PubMed Google Scholar -
Logan
S.,
Arzua
T.,
Canfield
S.G.,
Seminary
E.R.,
Sison
S.L.,
Ebert
A.D.,
Studying Human Neurological Disorders Using Induced Pluripotent Stem Cells: From 2D Monolayer to 3D Organoid and Blood Brain Barrier Models. Comprehensive Physiology.
2019;
9
(2)
:
565-611
.
View Article PubMed Google Scholar -
Boonekamp
K.E.,
Dayton
T.L.,
Clevers
H.,
Intestinal organoids as tools for enriching and studying specific and rare cell types: advances and future directions. Journal of Molecular Cell Biology.
2020;
12
(8)
:
562-8
.
View Article PubMed Google Scholar -
Bolla
S.R.,
Odeluga
N.,
Jetti
R.,
Histology, Bladder. StatPearls Publishing; Treasure Island, FL, USA: 2021. Histology, Bladder. 2021
.
-
Rossi
G.,
Manfrin
A.,
Lutolf
M.P.,
Progress and potential in organoid research. Nature Reviews. Genetics.
2018;
19
(11)
:
671-87
.
View Article PubMed Google Scholar -
Sobhani
A.,
Khanlarkhani
N.,
Baazm
M.,
Mohammadzadeh
F.,
Najafi
A.,
Mehdinejadiani
S.,
Multipotent Stem Cell and Current Application. Acta Medica Iranica.
2017;
55
(1)
:
6-23
.
PubMed Google Scholar -
Bartfeld
S.,
Clevers
H.,
Stem cell-derived organoids and their application for medical research and patient treatment. Journal of Molecular Medicine (Berlin, Germany).
2017;
95
(7)
:
729-38
.
View Article PubMed Google Scholar -
Kilani
R.T.,
Tamimi
Y.,
Hanel
E.G.,
Wong
K.K.,
Karmali
S.,
Lee
P.W.,
Selective reovirus killing of bladder cancer in a co-culture spheroid model. Virus Research.
2003;
93
(1)
:
1-12
.
View Article PubMed Google Scholar -
Galluzzi
L.,
Senovilla
L.,
Vitale
I.,
Michels
J.,
Martins
I.,
Kepp
O.,
Molecular mechanisms of cisplatin resistance. Oncogene.
2012;
31
(15)
:
1869-83
.
View Article PubMed Google Scholar -
Rudnicki
M.A.,
Cell culture methods and induction of differentiation of embryonal carcinoma cell lines. In Teratocarcinomas and Embryonic Stem Cells. A Practical Approach. 1987.;19–49. 1987
.
-
Ringuette Goulet
C.,
Bernard
G.,
Chabaud
S.,
Couture
A.,
Langlois
A.,
Neveu
B.,
Tissue-engineered human 3D model of bladder cancer for invasion study and drug discovery. Biomaterials.
2017;
145
:
233-41
.
View Article PubMed Google Scholar -
Sjöström
A.,
Bue
P.,
Malmstrom
P.,
Carlsson
J.,
Binding of 125I after administration of 125I-EGF-dextran, 125I-EGF or 125I to human bladder cancer spheroids. International Journal of Oncology.
2000;
17
(3)
:
559-64
.
View Article PubMed Google Scholar -
Bodgi
L.,
Bahmad
H.F.,
Araji
T.,
Al Choboq
J.,
Bou-Gharios
J.,
Cheaito
K.,
Assessing radiosensitivity of bladder cancer in vitro: A 2D vs. 3D approach. Frontiers in Oncology.
2019;
9
(MAR)
:
153
.
View Article PubMed Google Scholar -
Stellavato
A.,
Pirozzi
A.V.,
Diana
P.,
Reale
S.,
Vassallo
V.,
Fusco
A.,
Hyaluronic acid and chondroitin sulfate, alone or in combination, efficiently counteract induced bladder cell damage and inflammation. PLoS One.
2019;
14
(6)
:
e0218475
.
View Article PubMed Google Scholar -
Kim
H.,
Lee
S.J.,
Lee
I.K.,
Min
S.C.,
Sung
H.H.,
Jeong
B.C.,
Synergistic effects of combination therapy with akt and mtor inhibitors on bladder cancer cells. International Journal of Molecular Sciences.
2020;
21
(8)
:
2825
.
View Article PubMed Google Scholar -
Smith
Y.C.,
Grande
K.K.,
Rasmussen
S.B.,
O'Brien
A.D.,
Novel three-dimensional organoid model for evaluation of the interaction of uropathogenic Escherichia coli with terminally differentiated human urothelial cells. Infection and Immunity.
2006;
74
(1)
:
750-7
.
View Article PubMed Google Scholar -
Tannock
I.F.,
Lee
C.M.,
Tunggal
J.K.,
Cowan
D.S.,
Egorin
M.J.,
Limited penetration of anticancer drugs through tumor tissue: a potential cause of resistance of solid tumors to chemotherapy. Clinical Cancer Research.
2002;
8
(3)
:
878-84
.
PubMed Google Scholar -
Vatne
V.,
Litlekalsoey
J.,
Wentzel-Larsen
T.,
Hostmark
J.,
The in vitro effect of paclitaxel on a LacZ-transfected malignant transitional cell line. Anticancer research.
2005;
25
(3B)
:
2097-104
.
PubMed Google Scholar -
Warrick
J.I.,
Walter
V.,
Yamashita
H.,
Chung
E.,
Shuman
L.,
Amponsa
V.O.,
FOXA1, GATA3 and PPARIγ Cooperate to drive luminal subtype in bladder cancer: A molecular analysis of established human cell lines. Scientific Reports.
2016;
6
(1)
:
38531
.
View Article PubMed Google Scholar -
Varley
C.L.,
Southgate
J.,
Organotypic and 3D reconstructed cultures of the human bladder and urinary tract. Methods in Molecular Biology (Clifton, N.J.).
2011;
695
:
197-211
.
View Article PubMed Google Scholar -
Horsley
H.,
Dharmasena
D.,
Malone-Lee
J.,
Rohn
J.L.,
A urine-dependent human urothelial organoid offers a potential alternative to rodent models of infection. Scientific reports.
2018;
8
(1)
:
1238
.
View Article Google Scholar -
Horsley
H.,
Owen
J.,
Browning
R.,
Carugo
D.,
Malone-Lee
J.,
Stride
E.,
Ultrasound-activated microbubbles as a novel intracellular drug delivery system for urinary tract infection. Journal of Controlled Release.
2019;
301
:
166-75
.
View Article PubMed Google Scholar -
A.T. Hubbard,
N.V. Jafari,
N. Feasey,
J.L. Rohn,
A.P. Roberts,
Effect of Environment on the Evolutionary Trajectories and Growth Characteristics of Antibiotic-Resistant Escherichia coli Mutants. Frontiers in Microbiology.
2019;
10
:
2001
.
View Article Google Scholar -
Hoffmann
M.J.,
Koutsogiannouli
E.,
Skowron
M.A.,
Pinkerneil
M.,
Niegisch
G.,
Brandt
A.,
The new immortalized uroepithelial cell line HBLAK contains defined genetic aberrations typical of early stage urothelial tumors. Bladder Cancer (Amsterdam, Netherlands).
2016;
2
(4)
:
449-63
.
View Article PubMed Google Scholar -
Lee
S.H.,
Hu
W.,
Matulay
J.T.,
Silva
M.V.,
Owczarek
T.B.,
Kim
K.,
Tumor Evolution and Drug Response in Patient-Derived Organoid Models of Bladder Cancer. Cell.
2018;
173
(2)
:
515-528.e17
.
View Article PubMed Google Scholar -
Mullenders
J.,
de Jongh
E.,
Brousali
A.,
Roosen
M.,
Blom
J.P.,
Begthel
H.,
Mouse and human urothelial cancer organoids: A tool for bladder cancer research. Proceedings of the National Academy of Sciences of the United States of America.
2019;
116
(10)
:
4567-74
.
View Article PubMed Google Scholar -
Jana
S.,
Deo
R.,
Hough
R.P.,
Liu
Y.,
Horn
J.L.,
Wright
J.L.,
mRNA translation is a therapeutic vulnerability necessary for bladder epithelial transformation. JCI Insight.
2021;
6
(11)
:
1-12
.
View Article PubMed Google Scholar -
Whyard
T.,
Liu
J.,
Darras
F.S.,
Waltzer
W.C.,
Romanov
V.,
Organoid model of urothelial cancer: establishment and applications for bladder cancer research. BioTechniques.
2020;
69
(3)
:
193-9
.
View Article PubMed Google Scholar -
Yu
L.,
Li
Z.,
Mei
H.,
Li
W.,
Chen
D.,
Liu
L.,
Patient-derived organoids of bladder cancer recapitulate antigen expression profiles and serve as a personal evaluation model for CAR-T cells in vitro. Clinical & Translational Immunology.
2021;
10
(2)
:
e1248
.
View Article PubMed Google Scholar -
Yoshida
T.,
Okuyama
H.,
Nakayama
M.,
Endo
H.,
Nonomura
N.,
Nishimura
K.,
High-dose chemotherapeutics of intravesical chemotherapy rapidly induce mitochondrial dysfunction in bladder cancer-derived spheroids. Cancer Science.
2015;
106
(1)
:
69-77
.
View Article PubMed Google Scholar -
Santos
C.P.,
Lapi
E.,
Villarreal
J. Martinez de,
Alvaro-Espinosa
L.,
Fernandez-Barral
A.,
Barbachano
A.,
Urothelial organoids originating from Cd49f(high) mouse stem cells display Notch-dependent differentiation capacity. Nature communications.
2019;
10
(1)
:
4407
.
View Article Google Scholar -
Votanopoulos
K.I.,
Forsythe
S.,
Sivakumar
H.,
Mazzocchi
A.,
Aleman
J.,
Miller
L.,
Model of Patient-Specific Immune-Enhanced Organoids for Immunotherapy Screening: feasibility Study. Annals of Surgical Oncology.
2020;
27
(6)
:
1956-67
.
View Article PubMed Google Scholar -
Neal
J.T.,
Li
X.,
Zhu
J.,
Giangarra
V.,
Grzeskowiak
C.L.,
Ju
J.,
Organoid Modeling of the Tumor Immune Microenvironment. Cell.
2018;
175
(7)
:
1972-1988.e16
.
View Article PubMed Google Scholar -
Saini
G.,
Segaran
N.,
Mayer
J.L.,
Saini
A.,
Albadawi
H.,
Oklu
R.,
Applications of 3D bioprinting in tissue engineering and regenerative medicine. Journal of Clinical Medicine.
2021;
10
(21)
:
4966
.
View Article PubMed Google Scholar -
Chen
Y.C.,
Zhang
Z.,
Yoon
E.,
Yoon
E.,
Yoon
E.,
Yoon
E.,
Early Prediction of Single-Cell Derived Sphere Formation Rate Using Convolutional Neural Network Image Analysis. Analytical Chemistry.
2020;
92
(11)
:
7717-24
.
View Article PubMed Google Scholar -
Wu
K.M.,
A new classification of prodrugs: regulatory perspectives. Pharmaceuticals (Basel, Switzerland).
2009;
2
(3)
:
77-81
.
View Article PubMed Google Scholar -
Tatosian
D.A.,
Shuler
M.L.,
A novel system for evaluation of drug mixtures for potential efficacy in treating multidrug resistant cancers. Biotechnology and Bioengineering.
2009;
103
(1)
:
187-98
.
View Article PubMed Google Scholar -
Vunjak-Novakovic
G.,
Bhatia
S.,
Chen
C.,
Hirschi
K.,
HeLiVa platform: integrated heart-liver-vascular systems for drug testing in human health and disease. Stem Cell Research & Therapy.
2013;
4
(Suppl 1)
:
8
.
View Article PubMed Google Scholar -
Imura
Y.,
Sato
K.,
Yoshimura
E.,
Micro total bioassay system for ingested substances: assessment of intestinal absorption, hepatic metabolism, and bioactivity. Analytical Chemistry.
2010;
82
(24)
:
9983-8
.
View Article PubMed Google Scholar -
Drewitz
M.,
Helbling
M.,
Fried
N.,
Bieri
M.,
Moritz
W.,
Lichtenberg
J.,
Towards automated production and drug sensitivity testing using scaffold-free spherical tumor microtissues. Biotechnology Journal.
2011;
6
(12)
:
1488-96
.
View Article PubMed Google Scholar -
Chaw
K.C.,
Manimaran
M.,
Tay
E.H.,
Swaminathan
S.,
Multi-step microfluidic device for studying cancer metastasis. Lab on a Chip.
2007;
7
(8)
:
1041-7
.
View Article PubMed Google Scholar -
Zhang
C.,
Zhao
Z.,
Abdul Rahim
N.A.,
van Noort
D.,
Yu
H.,
Towards a human-on-chip: culturing multiple cell types on a chip with compartmentalized microenvironments. Lab on a Chip.
2009;
9
(22)
:
3185-92
.
View Article PubMed Google Scholar -
Kashaninejad
N.,
Nikmaneshi
M.R.,
Moghadas
H.,
Kiyoumarsi Oskouei
A.,
Rismanian
M.,
Barisam
M.,
Organ-Tumor-on-a-Chip for Chemosensitivity Assay: A Critical Review. Micromachines.
2016;
7
(8)
:
130
.
View Article PubMed Google Scholar -
Lanza
R.,
Langer
R.,
Vacanti
J.P.,
Atala
A.,
Principles of tissue engineeringAcademic press 2020.
Google Scholar -
Gosline
J.,
Lillie
M.,
Carrington
E.,
Guerette
P.,
Ortlepp
C.,
Savage
K.,
Elastic proteins: biological roles and mechanical properties. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences.
2002;
357
(1418)
:
121-32
.
View Article PubMed Google Scholar -
Lendlein
A.,
Kelch
S.,
Degradable, Multifunctional Polymeric Biomaterials with Shape-Memory. Materials Science Forum.
2005;
492\493
:
219-24
.
View Article Google Scholar -
Chen
Q.Z.,
Bismarck
A.,
Hansen
U.,
Junaid
S.,
Tran
M.Q.,
Harding
S.E.,
Characterisation of a soft elastomer poly(glycerol sebacate) designed to match the mechanical properties of myocardial tissue. Biomaterials.
2008;
29
(1)
:
47-57
.
View Article PubMed Google Scholar -
Chen
Q.Z.,
Harding
S.E.,
Ali
N.N.,
Lyon
A.R.,
Boccaccini
A.R.,
Biomaterials in cardiac tissue engineering: ten years of research survey. Vol. 59. Materials Science and Engineering R Reports.
2008;
59
(1-6)
:
1-37
.
View Article Google Scholar
Comments
Downloads
Article Details
Volume & Issue : Vol 11 No 10 (2024)
Page No.: 6801-6812
Published on: 2024-10-31
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 1925 times
- PDF downloaded - 636 times
- Supplement downloaded - 468 times
- XML downloaded - 62 times
