

Potential Protection of Mesenchymal Stem Cells Against Oxidative Stress With Hypoxia Preconditioning and Lipopolysaccharide Exposure
-
Fadhilla Maulany El Fajri

-
Vitriyanna Mutiara Yuhendri
-
Nicko Pisceski Kusika Saputra
-
Arfianti Arfianti
Copyrights: Fadhilla Maulany El Fajri, Vitriyanna Mutiara Yuhendri, Elmi Elmi, Nicko Pisceski Kusika Saputra, Arfianti Arfianti, 2025. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Background: Mesenchymal stem cells (MSCs) are among the most promising therapeutic options for degenerative diseases. However, the low viability and prolonged doubling time of MSCs during ex vivo expansion remain obstacles in their application in MSC-based therapies. MSC preconditioning has been considered a potential strategy to overcome challenges in MSC culture and to increase the therapeutic effects of MSCs. To mimic an inflammatory microenvironment and enhance stress resilience, lipopolysaccharide (LPS) was combined with hypoxia preconditioning. The aim of this study was to examine the effects of hypoxia and LPS preconditioning on viability, population doubling time, and expression levels of antioxidant genes in MSCs.
Methods: MSCs were isolated from Wharton’s jelly of the human umbilical cord tissue. Hypoxia was simulated using 100 μM CoCl₂, a chemical hypoxia mimetic, and cells were supplemented with 10 ng/mL LPS. To test oxidative stress resilience, preconditioned MSCs were exposed to 100 μM H2O2 for 24 hours. Cell viability was assessed using the CCK-8 assay, and population doubling time was calculated from cell counts obtained via trypan blue exclusion assays at 24-hour intervals. The mRNA expression levels of superoxide dismutase 1 (SOD1), catalase (CAT), and hypoxia-inducible factor 1α (HIF1α) were quantified by quantitative reverse transcription polymerase chain reaction (qRT-PCR). MSCs were divided into four groups: (1) normoxia (control), (2) hypoxia (CoCl₂), (3) LPS (10 ng/mL), and (4) hypoxia + LPS. Statistical analysis was performed using one-way ANOVA followed by Tukey’s post-hoc test.
Results: The results showed that the hypoxia + LPS preconditioning group significantly increased MSC viability compared with the hypoxia-alone group (p < 0.05). Similarly, MSCs in the hypoxia + LPS group exhibited a shorter population doubling time than untreated controls (p < 0.05). Preconditioned MSCs also demonstrated increased resistance to H₂O₂-induced oxidative stress, with higher viability compared to non-preconditioned cells (p < 0.05).
Conclusion: MSC preconditioning with hypoxia and LPS enhanced cell viability, proliferation capacity, and resistance to oxidative stress induced by H2O2. These findings suggest that combining hypoxia mimetics with inflammatory stimuli could optimize MSC culture conditions and improve therapeutic outcomes in degenerative diseases.
Introduction
Mesenchymal stem cells (MSCs) are multipotent stem cells that possess the ability to self-renew1, differentiate into various cell types, and exhibit immunomodulatory functions2. Owing to these properties, the development of MSCs as a cell-based therapy has gained significant interest. Numerous clinical trials have investigated their efficacy in treating conditions such as ischemic heart disease3, cardiovascular disorders4, liver fibrosis5, diabetes6, and bone injury7.
Despite their therapeutic potential, MSC-based therapy faces challenges. A major limitation is the low viability of MSCs under oxidative stress, which occurs when reactive oxygen species (ROS) exceed physiological levels. ROS can damage cell membranes, proteins, and DNA, leading to reduced MSC viability during in vitro expansion and compromising survival rates post-transplantation8. Strategies to mitigate oxidative stress are therefore critical for enhancing MSC viability, survival, and therapeutic outcomes.
Modification or preconditioning of MSC cultures is essential for optimizing therapeutic effects and maintaining viability during long-term culture9. Approaches include hypoxia preconditioning, exposure to inflammatory mediators (e.g., cytokines or lipopolysaccharides), and three-dimensional (3D) cell culture, all of which modulate MSC secretion of bioactive factors10, 11, 12.
Hypoxia, a key environmental factor, influences cellular processes such as metabolism, migration, proliferation, differentiation, and apoptosis13. Physiologically relevant hypoxic conditions (low oxygen tension) reduce ROS production, thereby minimizing oxidative damage that can trigger cell injury14. Short-term hypoxia (~5% O2) has been shown to enhance MSC proliferation and delay senescence15. For example, Arfianti et al. reported increased vascular endothelial growth factor (VEGF) secretion in hypoxia-preconditioned MSCs compared to normoxic cultures10. However, conflicting results, such as hypoxia-reoxygenation-induced cell death16, highlight the importance of methodological variations in mimicking oxygen stress.
Lipopolysaccharide (LPS), a Toll-like receptor 4 (TLR4) ligand derived from Gram-negative bacteria, modulates MSC function via TLR4 signaling12, 17, 18. Studies demonstrate that LPS preconditioning protects MSCs from oxidative stress-induced apoptosis and enhances survival through TLR4 and PI3K/Akt pathway activation12.
Methods
Isolation and Culture of MSC from Umbilical Cord
Mesenchymal stem cells in this study were isolated from Wharton’s Jelly (WJ) of umbilical cords obtained from four mothers who gave birth by elective cesarean section, with the following inclusion criteria: mothers aged 20-35 years, not in labor, no history of diabetes or hypertension during pregnancy, not infected with hepatitis B or HIV, no complications during pregnancy, and the fetus was not in fetal distress. The umbilical cord was placed in a pot containing phosphate-buffered saline (PBS) (Sigma-Aldrich) with 1% penicillin-streptomycin until the isolation process was performed. Umbilical cords obtained from cesarean sections had to be processed within 24 h. The umbilical cord was washed with PBS and cut into 5 cm segments. These segments were cut longitudinally, blood vessels were removed from the umbilical cord tissue, and WJ (perivascular tissue) was separated from the umbilical arteries and veins. Wharton’s jelly was chopped into ~1–2 mm pieces, transferred to a 75 cm² culture dish, and culture medium was added.
The culture medium consisted of Dulbecco’s modified Eagle’s medium (DMEM; Gibco) containing 5% human platelet lysate (HPL), 1% Glutamax, and 1% penicillin-streptomycin. The HPL used was pooled from three donors to avoid bias. The culture dish was incubated at 37°C in a CO2 incubator with 5% CO₂. After 10 days, spindle-shaped cell colonies appeared; the WJ pieces were then removed from the culture dish, and the culture medium was replaced every 3–4 days.
When the cell colonies reached 70–80% confluence, the cells were passaged using 0.25% trypsin-EDTA solution. Trypsin was inactivated using the MSC culture medium at a 1:1 (v/v) ratio. The cells were then centrifuged for 5 minutes at 1300 rpm at 4°C. They were passaged into a 75 cm² culture flask at a density of 3 × 10⁵ cells/mL. MSC characterization was performed by immunophenotyping using flow cytometry with a human mesenchymal stem cell analysis kit (BD Stemflow™). The criteria for MSC characterization were >95% of cells expressing CD73, CD90, and CD105 molecules and the ability to adhere to plastic (plastic-adherent).
Mesenchymal Stem Cell Culture with Hypoxic Conditions and LPS Exposure
MSC cultures at passages 3–4 that reached 70–80% confluence were prepared under hypoxic conditions. For viability testing, MSCs were grown on a 96-well plate at a density of 3 × 10⁴ cells/mL (five replicates per group) and allowed to attach for 24 h. Hypoxic conditions were induced using cobalt (II) chloride hexahydrate (CoCl₂, Sigma-Aldrich, Darmstadt, Germany). After MSCs reached ~80% confluence, the culture medium was replaced with fresh medium. For hypoxia induction, the culture medium was supplemented with 100 μM CoCl₂. For LPS exposure, the culture medium was supplemented with 10 ng/mL LPS (Sigma-Aldrich, St. Louis, MO, USA). The cells were then incubated for 24 h. Stock solutions of 25 mM CoCl₂ and 100 ng/mL LPS were prepared in sterile distilled water. The CoCl₂ dose was determined according to the study by Arfianti et al.10. To determine the LPS dose, a preliminary test was conducted in which MSCs were pretreated with different doses of LPS (0, 10, 100, and 1000 ng/mL) for 24 h, and viability was assessed. A dose of 10 ng/mL LPS was selected because it was the highest dose that did not cause cytotoxicity.
Induction of Oxidative Stress in MSC Cultures
After preconditioning with hypoxia, LPS, or both for 24 h, oxidative stress was induced in the cells by adding 100 μM H2O2 to the culture medium for 1 h.
MSC Viability Analysis
The effects of hypoxic conditions and LPS exposure on MSC viability were assessed using the Cell Counting Kit (CCK-8) assay. The CCK-8 assay was performed using WST-8 (2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,3-dimethoxyphenyl)-2H-tetrazolium chloride). WST-8 is a tetrazolium compound that is converted into a yellow formazan product by the dehydrogenase enzymes in living cells. After incubation, the amount of formazan produced was measured using a spectrophotometer at 450 nm. The intensity of the color correlates with the number of living cells: the more living cells, the stronger the yellow color and the higher the absorbance. The absorbance data were then compared with a standard to determine cell viability. After hypoxic culture and LPS exposure for 24 h, the culture medium was discarded, and 100 μL of serum-free medium plus 10 μL of CCK-8 reagent (Abbkine, Atlanta, Georgia, USA) were added to each well. The blank was a well without cells, used for comparison. The plates were incubated for 3 h, and absorbance was measured at 450 nm using a Multiskan SkyHigh microplate spectrophotometer (Thermo-Fisher Scientific). The percentage of viability was calculated using the following formula:
Doubling Time Analysis of MSC
The effects of hypoxic conditions and LPS exposure on MSC doubling time were assessed using the trypan blue exclusion assay. This method utilizes the principle that living cells will not be stained by trypan blue, whereas dead cells will absorb the dye. The cells were grown in 6-well plates until they reached 80% confluence and were then exposed to normoxia, hypoxia, LPS, or a combination of hypoxia and LPS. After 24 h, oxidative stress was induced by treating the cells with H2O2 for 1 h. MSCs were then detached by adding 0.25% trypsin-EDTA solution and incubating for 5 minutes in a CO2 incubator. Trypsin was inactivated by MSC culture medium at a 1:1 (v/v) ratio, and the cells were centrifuged at 1300 rpm for 5 minutes at 4°C. The supernatant was removed, and the cells were resuspended in 0.4% trypan blue (1:1 v/v). Cell counting was performed with an Improved Neubauer Hemocytometer under a light microscope at 10× magnification, recording the number of stained and unstained cells in five large squares (1 × 1 mm²). The doubling time was determined using the formula: Doubling time = Tx × (log(2) / (log(No) – log(Nx))) Where Tx is the culture duration (h), No is the number of cells at the beginning of treatment, and Nx is the number of cells at the end of treatment.
Gene Expression Analysis by Quantitative RT-PCR
The effects of hypoxic conditions and LPS exposure on the regulation of SOD1 and Catalase gene expression were analyzed by quantitative real-time polymerase chain reaction (qRT-PCR). After culturing under hypoxic conditions and LPS exposure, the culture medium was discarded, and total RNA was isolated from MSCs using the TRNzol Universal Reagent (Tiangen, Biotech, BJ, China). cDNA synthesis was performed by reverse transcription using 1 µg of RNA, followed by cDNA amplification with the GoTaq® 2-Step RT-qPCR System (Promega Corporation, Madison, WI, USA). A total of 1 μg of RNA was placed into a 0.2 mL RNase-free tube containing a mixture of dT primers, dNTPs, enzyme mix, and RNase-free H₂O. The tube was then incubated at 70°C for 5 min, 25°C for 10 min, 50°C for 50 min, and 85°C for 5 min. The resulting cDNA was diluted at a 1:50 ratio and stored at −20°C until further analysis. qRT-PCR was carried out using a Heal Force X 960 Real-Time PCR system with polymerase activation at 95°C, denaturation at 95°C, and annealing at 60°C.
Data Analysis
Numeric data are presented as mean ± standard deviation (SD). The effects of hypoxic conditions and LPS on MSC viability, doubling time, and SOD1 and catalase mRNA expression were analyzed by one-way ANOVA with post hoc Tukey’s multiple comparisons test. Statistical analyses were performed using GraphPad Prism v. 9. Differences were considered statistically significant at p < 0.05.
Results
Effect of Preconditioning with Hypoxia and LPS Exposure on MSC Viability
To analyze the effect of hypoxic conditions and LPS exposure on MSC viability, a CCK-8 assay was performed. Mesenchymal stem cells at passages 3–5 were seeded on 96-well culture plates, and the next day, MSCs were cultured under 1) normoxic conditions, 2) hypoxic conditions (100 µM CoCl₂), 3) exposure to 10 ng/mL LPS, and 4) a combination of hypoxia and LPS exposure. After 24 h, the CCK-8 assay was performed for 3 h. The absorbance was measured using a microplate reader at a wavelength of 450 nm.
Figure 1 shows the effects of preconditioning with hypoxic conditions and LPS exposure on MSC viability. MSCs cultured under a normoxic environment (control group) demonstrated a viability of 97.9%. Neither hypoxia nor LPS exposure alone for 24 h significantly altered MSC viability, with viabilities of 88.7% and 118.3%, respectively, when compared to the control group. However, the combination of hypoxia and LPS significantly enhanced MSC viability compared to the hypoxic group (p < 0.05).

Effect of Preconditioning with a Combination of Hypoxia and LPS Exposure on the Doubling Time of MSC
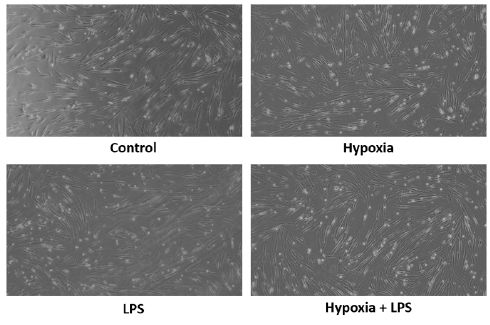
To measure the proliferation rates of MSCs after being cultured in hypoxic conditions and exposed to LPS, a doubling time analysis was performed using the trypan blue test. Figure 2 shows how preconditioning with hypoxia and LPS exposure affects the MSC doubling time. The time required for each group to double its population was as follows: the control group needed 688.8 h, the hypoxic group required 213.4 h, the LPS-treated group took 143.9 h, and the group exposed to both hypoxia and LPS took 71.13 h. These findings indicate that MSCs cultured under hypoxia and exposed to LPS had a significantly reduced doubling time compared to the control group (p < 0.05). Figure 2B compares MSC morphology following preconditioning for 24 h.

Effect of Preconditioning with a Combination of Hypoxia and LPS Exposure on the Gene Expression Levels of SOD1, Catalase, and HIF1-α
The expression of SOD1, catalase, and HIF1-α genes in MSCs was assessed following preconditioning under hypoxic conditions and LPS exposure, using qRT-PCR for mRNA expression analysis (Figure 3). Gene expression levels were normalized to GAPDH, which served as the reference control. The expression levels of SOD1 did not differ significantly among the experimental groups, although a slight increase was observed in MSCs exposed to LPS and the combination of hypoxia and LPS compared to the control. Similarly, catalase mRNA expression exhibited a modest increase in the hypoxic, LPS, and combination groups; however, these changes were not statistically significant. As a critical regulator of cellular adaptation to low oxygen levels, HIF1-α expression was expected to increase under hypoxic conditions. Nevertheless, the changes observed in the hypoxic group and the combination of hypoxia and LPS group were minimal and not statistically significant.

Discussion
The survival and proliferation of MSCs are highly dependent on the environmental conditions under which they are grown. Ideal conditions allow MSCs to achieve optimal efficacy in repairing damaged tissue after transplantation into patients. Low viability has been shown to affect therapy efficacy. The viability of transplanted cells after transplantation is influenced by various factors, including mechanical, cellular, and host factors19. One of the challenges in MSC-based therapy is the low viability and proliferation of MSCs due to oxidative stress8.
Preconditioning MSCs has been shown to improve MSC viability, which is affected by oxidative stress. Previous research has indicated that short-term hypoxic conditions, with an O2 concentration of approximately 5%, can stimulate cell proliferation without negatively impacting MSC survival15. Additionally, other studies have found that MSCs preconditioned with LPS are protected from oxidative stress-induced apoptosis12. However, no studies have yet explored the combination of hypoxic preconditioning and LPS on MSCs, nor have they examined their effects on oxidative stress and antioxidant activity. We expected that preconditioning MSC cultures would yield optimal therapeutic effects while preserving viability and promoting proliferation over extended culture periods.
We found that the combination of hypoxic conditions and LPS exposure significantly enhanced MSC viability (118.3%) compared with the group exposed solely to hypoxic conditions (88.77%). This is consistent with a study by Wang et al., which demonstrated that proper treatment with LPS can protect MSCs from apoptosis caused by oxidative stress and enhance their survival12.
Additionally, we found that MSCs cultured under hypoxic conditions and exposed to LPS exhibited a shorter doubling time than the control group, which was not subjected to either hypoxic conditions or LPS exposure. In line with this, a study by He et al. found that TLR4 activation by LPS enhanced the proliferation of MSCs in vitro20. In contrast, Antebi et al. found that MSCs proliferated considerably faster after 48 h under hypoxic conditions compared to those cultured under normoxic conditions. However, after 10 days of treatment, the MSCs in the hypoxic group also showed significantly slower proliferation21.
The expression of SOD1, catalase, and HIF1-α was higher in MSCs treated with hypoxic conditions and LPS than in the control group; however, these differences were not statistically significant. Similarly, Lavrentieva et al. also found significantly elevated HIF-1α protein levels in MSC cultures under hypoxic conditions, compared to barely detectable HIF-1α protein levels in the normoxic MSC population22.
SOD1 and catalase are controlled by the nuclear factor erythroid 2-related factor (NRF2), which serves as a transcriptional regulator of the cellular response to oxidative stress. NRF2 regulates antioxidant gene expression, and its activity is inhibited by Kelch-like ECH-associated protein (KEAP1), which binds to NRF2 in the cytosol and acts as a substrate adaptor. Under oxidative stress, a conformational change in KEAP1 prevents it from mediating NRF2 ubiquitination, leading to NRF2 translocation to the nucleus and activation of genes responsible for producing cytoprotective enzymes that detoxify ROS and other oxidants23.
The PI3K/Akt signaling pathway is often considered a key regulator of NRF2 activation. Duarte et al. demonstrated that the activation of PI3K/Akt promotes the translocation of NRF2 to the nucleus. The importance of PI3K/Akt signaling was emphasized in experiments involving LY294002, a PI3K inhibitor that significantly reduced NRF2 expression. This suggests that NRF2 is activated via two primary mechanisms. The first involves direct interaction with the NRF2/KEAP1 complex, causing a conformational change in KEAP1 that leads to its dissociation. The second mechanism relies on the activation of kinases, including PI3K/Akt and MAPKs23, 24, 25.
A previous study demonstrated that LPS can protect MSCs by activating TLR4 on their surface, thereby initiating the PI3K/Akt signaling pathway12. Furthermore, the Akt protein responds to oxygen levels in the body through hydroxylation. Under normoxic conditions, Akt activity may be inhibited by hydroxylation, whereas under hypoxic conditions, Akt is activated and plays a critical role in cell growth and survival26. This implies that a combination of hypoxic conditions and LPS exposure may represent an effective strategy for enhancing MSC survival. Activation of the PI3K/Akt pathway by the LPS-TLR4 interaction and hypoxia is anticipated to activate the NRF2 pathway, thereby promoting the expression of antioxidant genes. These findings underscore the need for further investigation of other potential mechanisms, beyond NRF2, which may contribute to this process.
We believe that the findings of this study can provide valuable insights into the development of methods to enhance the survival of MSCs under oxidative stress conditions, both during in vitro expansion and transplantation. However, this requires in vivo validation and further investigations. The limitation of this study was the use of CoCl₂ to simulate hypoxia. Future research should consider alternative methods, such as low-oxygen chambers, which provide a more stable and ideal environment for a more accurate replication of physiological hypoxia.
Conclusion
In this study, we highlighted the importance of preconditioning strategies for optimizing MSC function. This is especially important in MSC-based therapies, where cells often face harsh environments, including oxidative stress conditions, after transplantation or during in vitro culture.
In conclusion, the combination of hypoxic preconditioning and LPS exposure significantly enhances the viability and proliferation of MSCs under oxidative stress conditions, offering a promising strategy for improving the efficacy of MSC-based therapies. Our findings indicated that MSCs cultured under hypoxic conditions and treated with LPS showed a notable improvement in viability compared to hypoxic-only cultures. Additionally, MSCs subjected to combined hypoxic and LPS treatment demonstrated shorter doubling times, indicating enhanced proliferation.
Although the expression levels of antioxidant markers such as SOD1, catalase, and HIF1-α were elevated in the combined treatment group, these differences were not statistically significant, suggesting that other mechanisms may also contribute to the observed effects. These results highlight the potential of using hypoxia and LPS as preconditioning strategies to optimize MSC function, improve survival, and promote sustained proliferation under oxidative stress conditions, which is crucial for their therapeutic potential in tissue repair. Further studies are needed to explore the underlying molecular mechanisms and to assess the long-term effects of this combined preconditioning approach.
Abbreviations
3D – Three-dimensional; Akt – Protein kinase B; ANOVA – Analysis of variance; CAT – Catalase; CCK-8 – Cell Counting Kit-8; CD73 / CD90 / CD105 – Cluster of differentiation 73 / 90 / 105; cDNA – Complementary DNA; CoCl₂ – Cobalt(II) chloride; dNTPs – Deoxynucleotide triphosphates; DMEM – Dulbecco’s modified Eagle’s medium; DNA – Deoxyribonucleic acid; EDTA – Ethylenediaminetetraacetic acid; GAPDH – Glyceraldehyde-3-phosphate dehydrogenase; H₂O₂ – Hydrogen peroxide; HIF1-α (HIF1α) – Hypoxia-inducible factor 1-alpha; HIV – Human immunodeficiency virus; HPL – Human platelet lysate; KEAP1 – Kelch-like ECH-associated protein 1; LPS – Lipopolysaccharide; MAPKs – Mitogen-activated protein kinases; mRNA – Messenger RNA; MSC(s) – Mesenchymal stem cell(s); NRF2 – Nuclear factor erythroid 2-related factor 2; PBS – Phosphate-buffered saline; PI3K – Phosphatidylinositol 3-kinase; qRT-PCR – Quantitative reverse transcription polymerase chain reaction; RNA – Ribonucleic acid; ROS – Reactive oxygen species; SD – Standard deviation; SOD1 – Superoxide dismutase 1; TLR4 – Toll-like receptor 4; VEGF – Vascular endothelial growth factor; WJ – Wharton’s jelly; WST-8 – 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,3-dimethoxyphenyl)-2H-tetrazolium chloride.
Acknowledgments
We would like to thank the medical staff of the Department of Obstetrics and Gynecology, Hermina Hospital, for their valuable contribution in obtaining umbilical cord samples.
Author’s contributions
Arfianti Arfianti: Study conceptualization, methodology, validation, obtained funding, study supervision, data curation, software, intellectual input, writing – original draft preparation, review and editing. Fadhilla Maulany El Fajri: Software, analysis and interpretation of data, writing – original draft preparation, review and editing. Vitriyanna Mutiara Yuhendri: Software, analysis and interpretation of data, writing – original draft preparation. Elmi Elmi: Intellectual input and writing – original draft preparation. Nicko Pisceski Kusika Saputra: Resources and writing – original draft preparation. All authors read and approved the final manuscript.
Funding
This study was supported by Ministry of Education, Culture, Research, and Technology, Directorate of Research, Technology, and Community Indonesia 2024 (Contract no: 20588/UN19.5.1.3/AL.04/2024 and 20712/UN19.5.1.3/AL.04/2024).
Availability of data and materials
Data and materials used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
This research was approved by the Ethical Review Board for Medicine & Health Research, Faculty of Medicine, Universitas Riau (Ethical approval o: B/047/UN19.5.1.1.8/UEPKK/2021_Adendum3).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
-
Zhuang
W.Z.,
Lin
Y.H.,
Su
L.J.,
Wu
M.S.,
Jeng
H.Y.,
Chang
H.C.,
Mesenchymal stem/stromal cell-based therapy: mechanism, systemic safety and biodistribution for precision clinical applications. Journal of Biomedical Science.
2021;
28
(1)
:
28
.
View Article PubMed Google Scholar -
Eom
Y.W.,
Shim
K.Y.,
Baik
S.K.,
Mesenchymal stem cell therapy for liver fibrosis. Korean Journal of Internal Medicine (Korean Association of Internal Medicine).
2015;
30
(5)
:
580-9
.
View Article PubMed Google Scholar -
Alhomrani
M.,
Correia
J.,
Zavou
M.,
Leaw
B.,
Kuk
N.,
Xu
R.,
The Human Amnion Epithelial Cell Secretome Decreases Hepatic Fibrosis in Mice with Chronic Liver Fibrosis. Frontiers in Pharmacology.
2017;
8
:
748
.
View Article PubMed Google Scholar -
Cai
M.,
Shen
R.,
Song
L.,
Lu
M.,
Wang
J.,
Zhao
S.,
Bone Marrow Mesenchymal Stem Cells (BM-MSCs) Improve Heart Function in Swine Myocardial Infarction Model through Paracrine Effects. Scientific Reports.
2016;
6
(6)
:
28250
.
View Article PubMed Google Scholar -
Pei
Z.,
Zeng
J.,
Song
Y.,
Gao
Y.,
Wu
R.,
Chen
Y.,
In vivo imaging to monitor differentiation and therapeutic effects of transplanted mesenchymal stem cells in myocardial infarction. Scientific Reports.
2017;
7
(1)
:
6296
.
View Article PubMed Google Scholar -
Li
Y.,
Liu
J.,
Liao
G.,
Zhang
J.,
Chen
Y.,
Li
L.,
Early intervention with mesenchymal stem cells prevents nephropathy in diabetic rats by ameliorating the inflammatory microenvironment. International Journal of Molecular Medicine.
2018;
41
(5)
:
2629-39
.
View Article PubMed Google Scholar -
Satué
M.,
Schüler
C.,
Ginner
N.,
Erben
R.G.,
Intra-articularly injected mesenchymal stem cells promote cartilage regeneration, but do not permanently engraft in distant organs. Scientific Reports.
2019;
9
(1)
:
10153
.
View Article PubMed Google Scholar -
Suwanmanee
G.,
Tantrawatpan
C.,
Kheolamai
P.,
Paraoan
L.,
Manochantr
S.,
Fucoxanthin diminishes oxidative stress damage in human placenta-derived mesenchymal stem cells through the PI3K/Akt/Nrf-2 pathway. Scientific Reports.
2023;
13
(1)
:
22974
.
View Article PubMed Google Scholar -
Teixeira
F.G.,
Salgado
A.J.,
Mesenchymal stem cells secretome: current trends and future challenges. Neural Regeneration Research.
2020;
15
(1)
:
75-7
.
View Article PubMed Google Scholar -
Arfianti
A.,
Ulfah
Hutabarat
L.S.,
Agnes Ivana
G.,
Budiarti
A.D.,
Sahara
N.S.,
Hipoxia modulates the secretion of growth factors of human umbilical cord-derived mesenchymal stem cells. Biomedicine (Taipei).
2023;
13
(3)
:
49-56
.
View Article PubMed Google Scholar -
Madrigal
M.,
Rao
K.S.,
Riordan
N.H.,
A review of therapeutic effects of mesenchymal stem cell secretions and induction of secretory modification by different culture methods. Journal of Translational Medicine.
2014;
12
(1)
:
260
.
View Article PubMed Google Scholar -
Wang
Z.J.,
Zhang
F.M.,
Wang
L.S.,
Yao
Y.W.,
Zhao
Q.,
Gao
X.,
Lipopolysaccharides can protect mesenchymal stem cells (MSCs) from oxidative stress-induced apoptosis and enhance proliferation of MSCs via Toll-like receptor(TLR)-4 and PI3K/Akt. Cell Biology International.
2009;
33
(6)
:
665-74
.
View Article PubMed Google Scholar -
Ejtehadifar
M.,
Shamsasenjan
K.,
Movassaghpour
A.,
Akbarzadehlaleh
P.,
Dehdilani
N.,
Abbasi
P.,
The Effect of Hypoxia on Mesenchymal Stem Cell Biology. Advanced Pharmaceutical Bulletin.
2015;
5
(2)
:
141-9
.
View Article PubMed Google Scholar -
Bertram
C.,
Hass
R.,
Cellular responses to reactive oxygen species-induced DNA damage and aging. Biological Chemistry.
2008;
389
(3)
:
211-20
.
View Article PubMed Google Scholar -
Ito
A.,
Aoyama
T.,
Yoshizawa
M.,
Nagai
M.,
Tajino
J.,
Yamaguchi
S.,
The effects of short-term hypoxia on human mesenchymal stem cell proliferation, viability and p16(INK4A) mRNA expression: investigation using a simple hypoxic culture system with a deoxidizing agent. Journal of Stem Cells & Regenerative Medicine.
2015;
11
(1)
:
25-31
.
View Article PubMed Google Scholar -
Chen
T.L.,
Zhu
G.L.,
Wang
J.A.,
Wang
Y.,
He
X.L.,
Jiang
J.,
Apoptosis of bone marrow mesenchymal stem cells caused by hypoxia/reoxygenation via multiple pathways. International Journal of Clinical and Experimental Medicine.
2014;
7
(12)
:
4686-97
.
PubMed Google Scholar -
Pevsner-Fischer
M.,
Morad
V.,
Cohen-Sfady
M.,
Rousso-Noori
L.,
Zanin-Zhorov
A.,
Cohen
S.,
Toll-like receptors and their ligands control mesenchymal stem cell functions. Blood.
2007;
109
(4)
:
1422-32
.
View Article PubMed Google Scholar -
Hou
Y.S.,
Liu
L.Y.,
Chai
J.K.,
Yu
Y.H.,
Duan
H.J.,
Hu
Q.,
Lipopolysaccharide pretreatment inhibits LPS-induced human umbilical cord mesenchymal stem cell apoptosis via upregulating the expression of cellular FLICE-inhibitory protein. Molecular Medicine Reports.
2015;
12
(2)
:
2521-8
.
View Article PubMed Google Scholar -
Baldari
S.,
Di Rocco
G.,
Piccoli
M.,
Pozzobon
M.,
Muraca
M.,
Toietta
G.,
Challenges and Strategies for Improving the Regenerative Effects of Mesenchymal Stromal Cell-Based Therapies. International Journal of Molecular Sciences.
2017;
18
(10)
:
2087
.
View Article PubMed Google Scholar -
He
X.,
Wang
H.,
Jin
T.,
Xu
Y.,
Mei
L.,
Yang
J.,
TLR4 Activation Promotes Bone Marrow MSC Proliferation and Osteogenic Differentiation via Wnt3a and Wnt5a Signaling. PLoS One.
2016;
11
(3)
:
e0149876
.
View Article PubMed Google Scholar -
Antebi
B.,
Rodriguez
L.A.,
Walker
K.P.,
Asher
A.M.,
Kamucheka
R.M.,
Alvarado
L.,
Short-term physiological hypoxia potentiates the therapeutic function of mesenchymal stem cells. Stem Cell Research & Therapy.
2018;
9
(1)
:
265
.
View Article PubMed Google Scholar -
Lavrentieva
A.,
Majore
I.,
Kasper
C.,
Hass
R.,
Effects of hypoxic culture conditions on umbilical cord-derived human mesenchymal stem cells. Cell Communication and Signaling : CCS.
2010;
8
(1)
:
18
.
View Article PubMed Google Scholar -
Ngo
V.,
Duennwald
M.L.,
Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants.
2022;
11
(12)
:
2345
.
View Article PubMed Google Scholar -
Yoon
J.H.,
Youn
K.,
Jun
M.,
Protective effect of sargahydroquinoic acid against Aβ25-35-evoked damage via PI3K/Akt mediated Nrf2 antioxidant defense system. Biomedicine & Pharmacotherapy.
2021;
144
:
112271
.
View Article PubMed Google Scholar -
Duarte
A.,
Silveira
G.G.,
Soave
D.F.,
Costa
J.P.,
Silva
A.R.,
The Role of the LY294002 - A Non-Selective Inhibitor of Phosphatidylinositol 3-Kinase (PI3K) Pathway- in Cell Survival and Proliferation in Cell Line SCC-25. Asian Pacific Journal of Cancer Prevention : APJCP.
2019;
20
(11)
:
3377-83
.
View Article PubMed Google Scholar -
Guo
J.,
Chakraborty
A.A.,
Liu
P.,
Gan
W.,
Zheng
X.,
Inuzuka
H.,
pVHL suppresses kinase activity of Akt in a proline-hydroxylation-dependent manner. Science.
2016;
353
(6302)
:
929-32
.
View Article PubMed Google Scholar
Comments
Article Details
Volume & Issue : Vol 12 No 4 (2025)
Page No.: 7277-7285
Published on: 2025-04-30
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 228 times
- PDF downloaded - 71 times
- XML downloaded - 25 times
